figur 4: struktur af øjet.Dette billede er taget fra Chirstie (ref. 4) og er ophavsretligt begrænset i henhold til den givne kilde (dvs.det er ikke forfatterens eget arbejde).
øjets struktur: det område i øjet (se figur 4), der registrerer lys og farve, kaldes nethinden. De to typer detektionscelle til stede, stænger og kegler, behandler information, der kommer gennem linsen og sender den ned i synsnerven til hjernen. Stangceller (afsom der er omkring 100 millioner) registrerer graden af lethed, der kommer ind iøje og deres følsomhed afhænger af mængden af Rhodopsin til stede, somer selv genereret i cellerne. Rhodopsin ødelægges imidlertid afblegning ved udsættelse for lys, og derfor fungerer stangceller kun i svagt lys asved høj belysning fører det reducerede niveau af dette lysfølsomme pigment til Avery lav følsomhed. Kegleceller (hvoraf der er omkring 3 millioner) er også følsomme over for lysniveauer, menbevare deres funktion op til høj belysning ved brug af pigmentet Iodopsin.Detektion af farve er en funktion af de tre typer kegleceller, der findes inden for nethinden: mellem dem dækker de det synlige spektrum. Dette skyldes, at hver type er følsom over for et andet bølgelængdeområde med maksimumsvarende til rød (lang), grøn (medium) eller blå (kort).
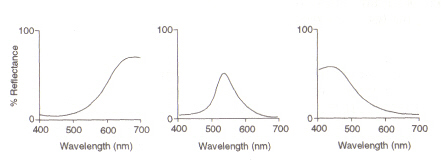
figur 5: maksimum på (fromvenstre) henholdsvis røde, grønne og blå kegleceller. Dette billede er taget fra Chirstie (ref. 4) og er ophavsretligt begrænset i henhold til den givne kilde (dvs.det er ikke forfatterens eget værk).
mekanismen for Rhodopsin i rodcellerne: proteinet Rhodopsin indeholder det protonatedretinal-Schiffs basiskompleks, som naturligt ligger i mellemmembranlommen dannet af de syv trans-membran a-helicalreceptorer. Der er mange flade skiver af rhodopsin inden for det ydre segment af arod-celle, som ved lysdetektion gennemgår en foto-isomer ændring fra Rhodopsin (11-cis) til all-trans retinal. Efter fotoisomeriseringskaskaden, der forekommer via 5 kortlivede intermediater (rutediagram 1), diffunderer trans retinal væk og omdannes tilbage til 11-cisretinal før genindtræden i cyklussen. Denne proces sker via reduktion toall-trans retinol efterfulgt af iltning/isomerisering i mørke. Fotoeksponeretrhodopsin (4.af de 5 mellemprodukter) udløser en kaskadeproces, der resulterer i hydrolyse af GMP. Dette lukker igen kationsspecifikke kanaler inden for stangcellemembranen, som naturligt er åben for tilstrømning af Na+i mørket, og på grund af effekten af hyperpolarisering sender den indre synatiske krop et nervesignal til andre neuroner i nethinden. Endelig lysinduceretsænkning af calciumniveauer hjælper genopretning af ophidsede neuroner til en passiv, “mørk”tilstand, og cyklussen starter igen ved detektering af lys. Detfotoreceptorer af kegleceller er også syv a-helicalreceptorer med 11-cis-retinal som deres chromophore. Detekteringsområdet varierer fra grønt til rødt, da de tre ikke-polære hydroksyl-holdige rester nearretinal erstattes sekventielt med polære.

venstre-figur 6: Position af 11-cis retinal inden for Rhodopsin. Dette billede er taget fra Stryer (ref. 6) og er ophavsretligt begrænset i henhold til den givne kilde (dvs.det er ikke forfatterens eget værk).
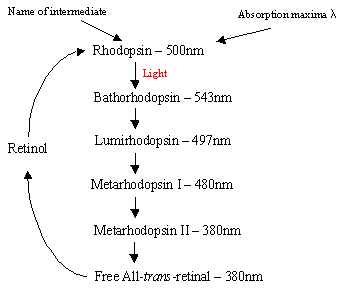
Højre Rutediagram 1: De 5 mellemprodukter i Rhodopsin-cyklussen.
hvor kommer Rhodopsin fra?
mennesker kan ikke lavehodopsin, i stedet bruger de og ekstern kilde, b-caroten, der findes i fødevarer for at syntese det:
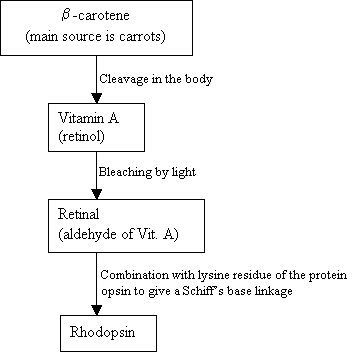 |
rutediagram 2: syntese af Rhodopsin nyt billede =figur 7: strukturerne af b-caroten (top), VitaminA (midten) og 11-cis retinal (bund). |
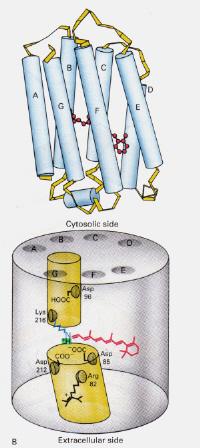
hvordan er mekanismen forbakterierhodopsin fundet i Halobakterier adskiller sig fra Rhodopsin fundet irodceller i det menneskelige øje?: Denne gang blokerer den protonerede retinal-Schiffs basekompleks naturligt en kanal gennem membranen, der ellers dannes af to tilstødende kamre. Det protonerede transkompleks donerer en proton til Asp – 85, som derefter tillader udgang af den samme proton til den ekstracellulære side. Fotoisomerisering til 13-cis-strukturen gør det muligt for Schiffs base at opsamleen proton fra Asp-96-remanensen på den cytosoliske side. Efter omlægning af cis-formen til trans fortsætter isomeriserings-og protonpumpecyklussen.