vi indsamlede frugter af L. barbarum og L. ruthenicum i fem udviklingsstadier, fra ung frugt (kur 10 dage efter blomstring) til moden (moden) frugt (34-45 dage efter blomstring) og studerede deres transkriptom og metabolom.
- RNA-sek de novo samling og funktionel annotation af unigener
- Lycium barbarum (LB): parvise interstage analyser
- LB transkriptom
- LB Metabolome
- Lycium ruthenicum (LR): parvise mellemtrinsanalyser
- LR-transkriptom
- LR-Metabolom
- interspecifik sammenlignende analyse af DEGs på forskellige udviklingsstadier
- Total DEGs under frugtudviklingen
- transkriptom-veje
- transkriptom – individuelle gener
- Metabolom – veje
- Metabolome-individuelle metabolitter
RNA-sek de novo samling og funktionel annotation af unigener
vi forberedte i alt 30 cDNA-biblioteker fra frugter af L. barbarum og L. ruthenicum, med tre biologiske replikater (tre frugter fra tre træer) på hvert tidspunkt: 2 arter, der er 5 tidspunkter, der er 3 biologiske replikater. Prøver blev mærket LB/LR(1-5)-(1-3), hvor LB er L. barbarum og LR er L. ruthenicum, 1-5 er udviklingsstadier af frugt (S1–S5), og 1-3 er individuelle prøver (biologiske replikater); så for eksempel repræsenterer LB1-1 L. barbarum, 1. prøveudtagede udviklingsstadium (S1), frugtprøve nr.1 (ud af tre). Vi genererede over 1.72 milliarder par-end læser for disse 30 cDNA biblioteker, svarende til et gennemsnit på 57.2 millioner læser pr prøve (supplerende datasæt S1). Streng kvalitetsvurdering og datafiltrering gav i alt 801.766 aflæsninger af høj kvalitet med den gennemsnitlige længde på 730 og N50 på 1107 bp (tabel 1). Endelig blev i alt 326.276 unigener med den gennemsnitlige længde på 596 bp og N50 på 847 bp opnået fra transkripterne (tabel 1). Korrelationskoefficienter for RNA-sekv-data for de 30 prøver indikerer meget god konsistens af resultater blandt biologiske replikater (Fig. 2).

Heatmap af korrelationskoefficienter for RNA-sek data for 30 prøver af L. barbarum (LB) og L. ruthenicum (LR) frugter på fem forskellige udviklingsstadier. Prøver er mærket LB / R1–5_1–3, Hvor LB er L. barbarum, LR er L. ruthenicum, 1-5 er frugt udviklingsstadier, og 1-3 individuelle prøver. Prøver blev grupperet efter hierarkisk klyngedannelse; dendrogrammer over og til venstre for varmekortet indikerer sammenhæng mellem prøver.
blandt alle 326.276 unigener, der blev spurgt mod offentlige databaser, blev i alt 193.021 (59,15%) matchede gener og/eller proteiner i mindst en database og 12.171 (3,73%) kommenteret i alle databaser. Det største antal unigener (149.863, 45,93%) blev kommenteret i NT-databasen og det laveste antal (24.017; 7,36%) i KOG-databasen.
Lycium barbarum (LB): parvise interstage analyser
LB transkriptom
det højeste antal grader blev identificeret i alle parvise sammenligninger af 1.trin og i 2. vs. 5. trin sammenligning (alle > 10.000 grader; tabel 2). De mindste tal blev identificeret i 3. vs. 4. og 4. vs. 5. trin sammenligninger (255-257). Heatmap-analyse af DEGs i LB viser, at temmelig forskellige sæt gener blev stærkt opreguleret i de tidlige udviklingsstadier (1 + 2) og i senere stadier (3 til 5) (Fig. 3A). Prøverelateret analyse indikerer, at prøverne kunne opdeles i to klader (trin 1 + 2 og 3 + 4 + 5), med sidstnævnte klade yderligere opdelt i to klader: trin 3 + 4 og trin 5. Intraspecifik KEGG funktionel klassificeringsanalyse af disse DEGs identificerede 15 veje signifikant (P < 0,05) differentielt reguleret mellem forskellige udviklingsstadier (supplerende datasæt S2). Særligt stærkt differentielt reguleret var ‘plantehormonsignaltransduktion’, ‘phenylpropanoid biosyntese (b.)’, ‘linolsyremetabolisme (m.)’, ‘ stivelse og saccharose m.( Fig. 3A).

Heatmaps og funktionelle vejanalyser af differentielt udtrykte gener (DEGs) i Lycium barbarum (panel A) og L. ruthenicum (panel B) frugter. Heatmaps blev genereret ved en hierarkisk analyse af DEGs (y-akse) og individuelle prøver (h-akse), hvor dendrogrammer over og til venstre for heatmap indikerer sammenhæng mellem prøver. Prøver er mærket LB / R_1–5_1-3, Hvor artsakronymet (LB eller LR) efterfølges af frugtens udviklingsstadium (1-5) og prøvenummeret (1-3). Intraspecifikke KEGG-vejanalyser af DEGs i alle fem udviklingsstadier i de to arter er vist til højre for heatmaps. Kun de top 15 berigede veje er angivet. s-værdien er en FDR-justeret p-værdi.
For en mere dybdegående analyse af dataene fokuserede vi på sammenligningen af de mest markant regulerede veje i successive udviklingsstadier. I den første parvise sammenligning (S1 vs S2) var ‘phenylpropanoid b.’ den mest differentielt regulerede vej efterfulgt af ‘stivelse og saccharose m.’ (Fig. 4). Meget stort antal DEGs (> 100) blev identificeret i begge veje. Et lignende resultat blev observeret i den følgende parvise sammenligning, S2 vs. S3, men på trods af det ret store antal DEGs (>80) ‘stivelse og saccharose m.’udstillet noget lavere K-værdi. I sammenligningen i S3 vs. S4 var ‘kulstoffiksering i fotosyntetiske organismer’ den mest markant regulerede vej, men antallet af gener var meget lavere. I det sidste par var S4 vs. S5, ‘seatin b.’, flavonoid b.’, fedtsyre b.’ og ‘galactose m.’ de mest markant regulerede veje, men ingen af stierne udviste mere end to grader.

lb transkriptom: parvis INTERSTAGE Kegg metabolisk vej berigelse. A) Fase 1 vs. fase 2. B) fase 2 vs. trin 3. C) fase 3 vs. fase 4. D) fase 4 vs. fase 5. Top 15 (eller alle hvis total <15) signifikant berigede veje vises. Barens farve repræsenterer størrelsen af K-værdien (en FDR-justeret p-værdi). Farvediagram er vist i figuren.
LB Metabolome
vi gennemførte parvise sammenligninger af udviklingsstadier for at identificere berigelsen af metabolitter mellem alle trinpar. Antallet af differentielt regulerede metabolitter i parvise sammenligninger af udviklingsstadier i LB varierede fra 66 (trin 3 vs 4) til 129 (trin 2 vs 5). Det største antal i successive trin sammenligninger blev observeret mellem 1.og 2. trin (117). Disse blev tildelt et stort antal veje; med det største antal metabolitter tildelt ‘b. af sekundære metabolitter’ (også den laveste p-værdi) efterfulgt af ‘proteinfordøjelse og absorption’ og ‘b.af aminosyrer’ (Fig. 5; Supplerende Tal). I den successive fase par sammenligning (S2 vs. S3),’ ABC transportører ‘var den mest signifikant beriget vej, efterfulgt af’purine m.’. I S3 vs. S4 par sammenligning, ‘mikrobiel m. i forskellige miljøer’ var den mest markant berigede vej, efterfulgt af ‘carbapenem m.’. I S4 vs. S5-parsammenligningen udviste et relativt stort antal veje lignende resultater (to metabolitter og lignende p-værdier), men bemærkelsesværdigt er udseendet af ‘isoflavonoid b.’ og ‘flavonoid b.’ blandt dem (Fig. 5; Supplerende Tal).

lb metabolomiske data: parvis INTERSTAGE Kegg metabolisk vej berigelse. A) Fase 1 vs. fase 2. B) fase 2 vs. fase 3. C) fase 3 vs. fase 4. D) fase 4 vs. fase 5. Rich factor er forholdet mellem antallet af signifikant regulerede metabolitter i vejen og det samlede antal metabolitter, der er kommenteret i den vej (interval = 0 til 1,0). Punktets størrelse repræsenterer antallet af signifikant berigede metabolitter i den tilsvarende vej, og punktets farve repræsenterer P-værdien (begge legender vist i figuren).
med hensyn til individuelle metabolitter blev listen over metabolitter, der blev opreguleret i S1, i S1 vs. S2-sammenligningen toppet af Trehalose, Glactinol og L-æblesyre (alle kar 20 log2FC). Oliesyre, 2-Oksoadipinsyre og stearinsyre var de mest opregulerede metabolitter i S2 (alle CRT 15 til 18 log2FC). Listen over de mest opregulerede metabolitter i S2 sammenlignet med S3 blev toppet af Dihydroksyaceton, LysoPC(18:1(9 gange)) og adenin (alle 16 til 17,5 log2fc). Listen over de mest opregulerede metabolitter i S3 (sammenlignet med S2) blev toppet af Trehalose, Galactinol og L-æblesyre (alle opregulerede 19 til 20 log2FC). I S3 vs. S4-sammenligningen blev listen over mest stærkt opregulerede metabolitter i S3 toppet af L-æblesyre, dl-arginin og oliesyre (alle kar 18 til 19 log2FC), mens listen over de mest opregulerede metabolitter i S4 blev toppet af D-Mannose, N-Acetyllactosamin og LysoPC(18:1(9 gange)) i LB (alle kar 16 log2FC). I S4 vs. S5-sammenligning blev listen over de mest opregulerede metabolitter i S4 toppet af Trehalose, palmitinsyre, N-Acetyllactosamin (alle liter 17 til 18 log2FC) i LB. Listen over de mest stærkt opregulerede metabolitter i S5 blev toppet af L-Norleucin, anthranilsyre (vitamin L1) og DL-arginin (alle liter 16 til 18 log2FC) (supplerende datasæt S3).
Lycium ruthenicum (LR): parvise mellemtrinsanalyser
LR-transkriptom
Heatmap-analyse af DEGs i LR viser, at næsten helt forskellige sæt gener var stærkt opreguleret i de tidlige udviklingsstadier (1 + 2) og i moden frugt (trin 5) med en tilsyneladende transkriptomisk nulstilling, der forekom efter anden fase (Fig. 3B). Prøverelateret analyse indikerer, at prøverne kunne opdeles i to klader (trin 1 + 2 og 3 + 4 + 5), med sidstnævnte klade yderligere opdelt i to klader: trin 3 + 4 og trin 5. Intraspecifik KEGG funktionel klassificeringsanalyse af disse DEGs identificerede 35 veje signifikant (P < 0,05) differentielt reguleret mellem forskellige udviklingsstadier (supplerende datasæt S2). Særligt stærkt differentielt reguleret var ‘fotosyntese’, ‘linolsyre m.’, ‘isokinolinalkaloid b.’, ‘flavonoid b.’ og ‘stivelse og saccharose m’ (Fig. 3B). Det højeste antal grader blev identificeret i de fleste parvise sammenligninger af trin 1 (1 vs. 2 var en undtagelse) og i trin 2 vs. 5 sammenligning (alle >5.000 grader; tabel 2). Langt det mindste antal blev identificeret i S3 vs. S4 sammenligning (39). I S1 vs S2 parvis sammenligning var’ aminosukker og nukleotidsukker m. ‘den mest differentielt regulerede vej efterfulgt af’ linolsyre m. ‘ (Fig. 6). I S2 vs. S3 parvis sammenligning var de mest signifikant differentielt regulerede veje ‘isokinolinalkaloid b.’, ‘tyrosin m.’, ‘phenylpropanoid b’og’ flavonoid b.’. I begge parvise sammenligninger blev det højeste antal DEGs (henholdsvis>15 og >35) identificeret i ‘stivelse og saccharose m.’. I S3 vs. S4 sammenligning, ‘ (alfa -) linolsyre m.’ var den mest signifikant regulerede vej, men antallet af gener var meget lavere. I S4 vs. S5-sammenligningen var ‘ phenylpropanoid b ‘(også det største antal DEGs) og’ linolsyre m. ‘ de mest signifikant regulerede veje.
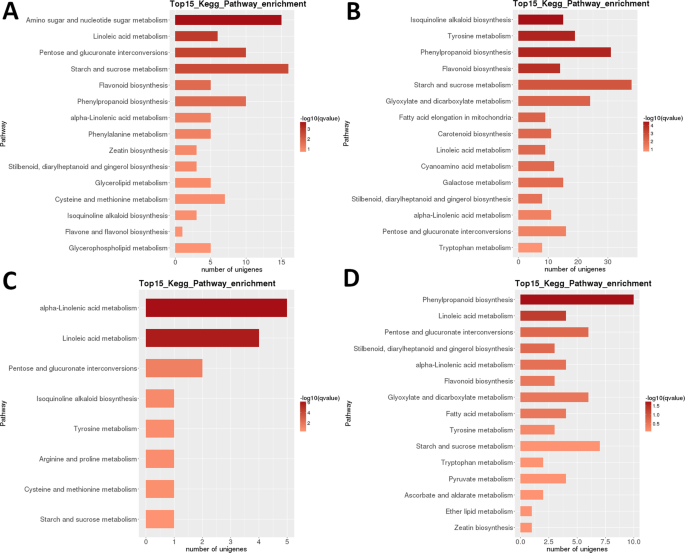
LR transkriptom: parvis INTERSTAGE Kegg metabolisk vej berigelse. A) Fase 1 vs. fase 2. B) fase 2 vs. fase 3. C) fase 3 vs. fase 4. D) fase 4 vs. fase 5. Top 15 (eller alle hvis total <15) signifikant berigede veje vises. Barens farve repræsenterer størrelsen af K-værdien (en FDR-justeret p-værdi, farvediagram vist i figuren).
LR-Metabolom
antallet af differentielt regulerede metabolitter i parvise sammenligninger af udviklingsstadier i LR varierede fra 66 (trin 3 vs 4) til 133 (trin 2 vs 5) (supplerende datasæt S3). I successive trin sammenligninger blev det største antal observeret mellem 1.og 2. trin (117). Disse blev tildelt et stort antal veje; med det største antal metabolitter tildelt ‘b af sekundære metabolitter ‘efterfulgt af’ proteinfordøjelse og absorption’,’ b. af aminosyrer ‘og’flavonoid b’. I S2 vs. S3-sammenligningen var ‘ABC-transportører ‘den mest signifikant berigede vej efterfulgt af’purine m’. I S3 vs. S4 par sammenligning, ‘mikrobiel m. i forskellige miljøer’ var den mest markant berigede vej, efterfulgt af ‘carbapenem m.’. I S4 vs. S5 par sammenligning, et relativt stort antal veje udviste lignende resultater (2 metabolitter og lignende p-værdier), men bemærkelsesværdigt er udseendet af ‘isoflavonoid b.’ og ‘flavonoid b.’ blandt dem (Fig. 7; Supplerende Tal).

LR-metabolomiske data: parvis INTERSTAGE Kegg metabolisk vej berigelse. A) Fase 1 vs. fase 2. B) fase 2 vs. fase 3. C) fase 3 vs. fase 4. D) fase 4 vs. fase 5. Rich factor er forholdet mellem antallet af signifikant regulerede metabolitter i vejen og det samlede antal metabolitter, der er kommenteret i den vej (interval = 0 til 1,0). Punktets størrelse repræsenterer antallet af signifikant berigede metabolitter i den tilsvarende vej, og punktets farve repræsenterer P-værdien (begge legender vist i figuren).
for så vidt angår individuelle metabolitter (supplerende datasæt S3), i S1 vs. S2-sammenligning blev listen over metabolitter, der blev opreguleret i S1, toppet af Trehalose, Galactinol og L-æblesyre (lut19-21 log2FC), mens oliesyre, 2-Oksoadipinsyre og stearinsyre var de mest opregulerede metabolitter i S2 (lut15-18 log2FC). I S2 vs. S3-sammenligningen var Dihydroksyaceton, indoksylsulfat og N-Acetyllactosamin de mest opregulerede metabolitter i s2 (Kurt 17-19, 5 log2FC) og Trehalose, Galactinol og L-æblesyre (Kurt 19-21 log2FC) i S3. I S3 vs. S4-sammenligning, L-æblesyre, dl-arginin og oliesyre blev opreguleret i S3 (Kurt 16-19 log2FC) og 1,7-Kurt, D-Mannose og N-Acetyllactosamin (Kurt 15-17 log2FC) i S4. I S4 vs. S5-sammenligningen blev Flavinmononukleotid, Trehalose og Isoferulinsyre opreguleret i S4 (kur 18-20 log2FC), hvorimod PG(16:0/18:1(9%)), D-prolin og DL-arginin var mest opregulerede (alle kur 16 til 18 log2FC) metabolitter.
interspecifik sammenlignende analyse af DEGs på forskellige udviklingsstadier
Total DEGs under frugtudviklingen
interspecifik parvis fase sammenligning (LR1 vs. LB1, LR2 vs. LB2 osv.) viser, at 928 grader blev delt af alle fem par (Fig. 8A). Det højeste antal DEGs blev identificeret i trin 3 (3989) og det laveste i trin 4 (2825) (Fig. 8B); der henviser til, at det højeste antal grader, der er unikke for et par, blev observeret i trin 3 (574), 1 og 5 (begge 554) og det laveste i trin 4 (126) (Fig. 8A). Antallet af op-og nedregulerede grader var relativt ens i hver af de parvise trin sammenligninger; f.eks. i trin 5 blev 1668 grader opreguleret, og 1670 grader blev nedreguleret i LR sammenlignet med LB (Fig. 8B). I de andre fire faser var antallet af opregulerede gener dog lidt (93 til 189 grader) højere.

gener differentielt udtrykt (DEGs) mellem frugter af L. barbarum (LB) og L. ruthenicum (LR). (A) detaljerede trinvis (1 til 5) sammenligninger (LB vs. LR). B) antallet af opregulerede (røde) og nedregulerede (grønne) grader i LR sammenlignet med LB i fem undersøgte udviklingsstadier.
transkriptom-veje
Heatmap analyse af DEGs indikerer, at frugterne af to arter udviser meget forskellige genekspressionsprofiler i alle udviklingsstadier, men biologiske replikater udviste meget lignende profiler, hvilket indikerer en begrænset mængde individuel variabilitet i hvert udviklingsstadium (supplerende figurer: Fig. S9). Sammenlignende analyse af Kegg-vejberigelse viser, at kun nogle veje konsekvent var stærkt beriget (med hensyn til genregulering) i LR sammenlignet med LB gennem alle fem udviklingsstadier (Fig. 9). Især transduktion af plantehormonsignal (2. højeste i S1, 8.højeste i S2, den højeste i S3, S4 og S5) og interaktion mellem plante og patogen (den højeste i S1, 3. højeste i S2, 2. højeste i S3, 4. højeste i S4 og 15. højeste i S5) var relativt højt opreguleret i alle faser. Phenylpropanoid biosyntese (ikke i top 15 I S1, den højeste i S2, 3.-højeste i S3, 7.-højeste i S4, 11.-højeste i S5), allestedsnærværende og anden terpenoid-kinonbiosyntese (ikke i top 15 i S1, 6.-højeste i S2, 7.-højeste i S3, 2.-højeste i S4, 6.-højeste i S5) var også relativt højt opreguleret i alle faser undtagen den første. Flavonoid biosyntesevej blev ikke stærkt beriget i tidlige stadier (ikke i top 15 I S1, 14.højeste i S2) og stærkt beriget i sene stadier (3. til 4. højeste i trin 3 til 5). (alfa-)Linolsyremetabolisme blev stærkt beriget i mellemtrin (9.-højeste i S1, 2.-højeste i S2, 5. og 6.-højeste i S3, 8.-højeste i S4, ikke i top 15 i S5).

sammenlignende analyse af Kegg metabolisk vej berigelse. De øverste 15 veje beriget i L. ruthenicum i sammenligning med L. barbarum er vist til venstre (rød), og dem beriget i L. barbarum i sammenligning med L. ruthenicum til højre (grøn). Udviklingsstadier (1-5) er angivet i figuren. s-værdien er en FDR-justeret p-værdi.
blandt de veje nedreguleret i L. ruthenicum i sammenligning med L. barbarum (Fig. 9) bemærkelsesværdige ændringer blev observeret mellem de tidlige stadier (1 og 2), Når cyanoaminosyremetabolisme og carotenoidbiosyntese var mest nedreguleret, og sene stadier (4 og 5), når SNARE interaktioner i vesikulær transport, nicotinat og nicotinamidmetabolisme og porfyrin og chlorophyll metabolisme var konsekvent relativt stærkt nedreguleret.
transkriptom – individuelle gener
blandt de mest differentielt udtrykte gener var nogle udviklingsstadiespecifikke (dvs.meget differentielt reguleret kun i tidlige eller sene udviklingsstadier), men nogle var konsekvent meget differentielt reguleret gennem alle fem undersøgte stadier (supplerende datasæt S4). Flere immunitetsrelaterede gener meget stærkt opreguleret i LR sammenlignet med LB i de tidlige udviklingsstadier er blandt eksemplerne på det udviklingsstadiumspecifikke ekspressionsmønster: chitinase var den 2. højeste opregulerede DEG i S1 (13,43 gange), den højeste i S2 (13,89 gange), men i senere faser var det ikke en DEG. 1/2 var også meget stærkt opreguleret i tidlige stadier, den højeste i S1 (13.70) og 2.højeste i S2 (10.90), men det blev heller ikke identificeret som DEG i senere stadier. Nogle vækstrelaterede gener udviste også et lignende ekspressionsmønster: phosphoglyceratkinase (PGK) var blandt de håndfuld mest opregulerede gener i de første tre faser (13.14, 12.87 og 12.77 henholdsvis), men det blev heller ikke identificeret som DEG i senere faser. CCR4-ikke transkriptionskompleks underenhed 7/8 (CNOT7/8) udviste også et meget lignende ekspressionsmønster: stærkt opreguleret i de første tre faser og ikke en DEG i trin 4 og 5. Flere flavonoid og phenylpropanoid biosynteseassocierede gener udviste et omvendt udviklingsstadiumspecifikt ekspressionsmønster med relativt lav ekspression i tidlige stadier og meget høj i senere stadier. Eksempler er: bifunktionel dihydroflavonol 4-reduktase / flavanon 4-reduktase (DFR), som var let opreguleret i LR i S1 (2.25), ikke en DEG i S2, stærkt opreguleret i S3 (7,79) og 3.højeste opregulerede gen i S4 (14,25) og S5 (16,03). En paralogue af dette gen udviste et næsten identisk mønster: Let opreguleret i S1 (2.44), ikke en DEG i S2, stærkt opreguleret i S3 (7.40), 6.-højeste opregulerede DEG i S4 (13.26) og 5. i S5 (14.59). Tilsvarende var flavonoid 3′, 5′-hydroksylase (F3 ‘ 5 ‘ H) ikke en DEG i de første to faser, stærkt opreguleret i S3 (6,69), 5.højeste opregulerede gen i S4 (13,42) og 4. højeste i S5 (15,05). Flavonoid O-methyltransferase (OMT) var ikke en DEG i S1, men i S2 udviste den allerede et mellemhøjt opreguleringsniveau (4.32), ved S3 var det allerede det tredje højeste opregulerede DEG (13.30), og det var det højest opregulerede gen i S4 (18.73) og S5 (18.10). Leucoanthocyanidindioksygenase (ldoks; anthocyaninbiosyntese) var ikke en DEG i S1 og S2, efterfulgt af høj til meget høj opregulering i senere stadier (henholdsvis 5,63, 9,44, 11,56). To chalcone synthase paraloger (CHS og CHS2; flavonoid biosyntese) var heller ikke stærkt reguleret i S1 og S2 (CHS2: ikke en DEG, CHS: -1.14 i S1, ikke en DEG i S2), men i S3–S5 udviste begge gener en mellemhøj til høj opregulering (CHS2: 5.32, 7.84, 6.00; og CHS: 4.67, 7.01, 6.82; henholdsvis). Vi valgte disse gener til kpcr-analyse, og resultaterne er meget kongruente med RNA-sekv-dataene (supplerende resultater; supplerende datasæt S5). Endelig blev cytokinindehydrogenase, et biosynteserelateret gen, også i stigende grad opreguleret i de sidste tre faser (2,6-5,7).
imidlertid blev nogle gener konsekvent forskelligt udtrykt gennem alle fem undersøgte stadier. Eksempler omfattede også nogle immunitetsrelaterede gener, såsom to paraloger af glutathion S-transferase, stærkt opreguleret i LR sammenlignet med LB i alle faser: 9.38 og 8.58 (alle værdier præsenteret som fold-ændringer i respektive rækkefølge) i S1, 6.30 og 6.34 i S2, 2.og 7. højeste opregulerede grader i S3 (14.08 og 12.70), 2. og 4. højeste i S4 (15.71 og 14.16) og 2. og 6. højeste i S5 (16.48 og 14.40). Plantesygdomsresistensprotein RPM1 blev også stærkt opreguleret i alle fem trin (S1 = 13,15; S2 = 12,08; S3 = 13,11, S4 = 12,81; S5 = 13:94). Blandt de konsekvent differentielt udtrykte gener gennem alle udviklingsstadier var også nogle relateret til aminosyremetabolismen, men deres mønster blev vendt i sammenligning med tidligere eksempler: de udviste høj nedregulering i LR sammenlignet med LB. Eksempler er acetyl-CoA acyltransferase 1 (aat1; valin, leucin og isoleucin nedbrydning), med en tidsmæssig profil af stadig højere nedregulering, startende fra -7,0 i S1 til <−10 gange i de sidste tre faser. Proliniminopeptidase, forbundet med arginin og prolinmetabolisme, blev stærkt nedreguleret i LR i alle faser: S1 = -9,75, S2 = -10,89 (3.-højeste), S3 = -11,05 (4.-højeste), S4 = -10,01 og S5 = -11,98 (3.-højeste). Endelig blev 5–methyltetrahydropteroyltriglutamat-homocysteinmethyltransferase (metE) konsekvent ekstremt stærkt nedreguleret i LR i alle faser: 2.-højeste i S1 (-11,76), den højeste i s2 (-11,74), 3.-højeste i S3 (-11,43), den højeste i S4 (-12,36) og 2.-højeste i S5 (-12,83). To DNA-replikation og transkriptionsassocierede gener blev også stærkt nedreguleret i LR i alle faser: GTP-bindende nukleart protein Ran (RAN; -10,0 til -12,0) og replikationsfaktor A1 (RFA1) (-8,0 til -12,0). Nogle vækst-og stressrelaterede gener blev også konsekvent stærkt nedreguleret i LR: heterogent nukleart ribonukleoprotein A1/A3 (hnRNP; -7 til -11) og varmechok 70 kDa protein 1/8 (HSPA1_8) S1 = -4,95, S2 = -8,88, S3 = -11,48 (2.højeste), S4 = -9,36, S5 = -12,89 (den højeste). Interessant nok blev et phenylpropanoid biosynteserelateret gen, shikimate hydroksycinnamoyltransferase (HCT), også konsekvent stærkt nedreguleret i LR: S1 = -6,82, S2 = -8,14, S3 = -11,71 (den højeste), S4 = -11,00 (3.-højeste), S5 = -11,91 (4.-højeste). Imidlertid blev en nøgleregulator for anthocyaninbiosyntese, transkriptionsfaktor MYB114, stærkt opreguleret i LR i alle fem udviklingsstadier: 6.11, 4.69, 7.47, 9.05 og 8.95 (henholdsvis S1–S5).
Metabolom – veje
vi gennemførte også en sammenlignende interspecifik trinvis analyse af metaboliske veje (Fig. 10). I det første udviklingsstadium (S1) identificerede vi 39 differentielt regulerede metabolitter. Blandt de 20 bedste veje, som disse metabolitter var forbundet med, var flere af dem forbundet med aminosyrer, men det samlede antal metabolitter pr.vej var relativt lille (1-2), og P-værdier antydede ikke et højt signifikansniveau (Fig. 10-panel 1). Især vitamin B6-metabolisme (m) og mikrobiel m i forskellige miljøer udviste begge relativt høje P-værdier, berigelsesfaktor (EF) på 1,0 og 2 identificerede metabolitter. I S2 identificerede vi 58 differentielt regulerede metabolitter, der kun var forbundet med fire veje: tryptophan m, phenylpropanoid biosyntese (b), B af phenylpropanoider (disse er to forskellige veje i KEGG-databasen) og phenylalanin, tyrosin og tryptophan b (alle EF = 1,0, 2-3 metabolitter og p < 0,5; Fig. 10-panel 2). I S3 identificerede vi 59 differentielt regulerede metabolitter, der var forbundet med 19 veje, mest Alle med EF 1.0, men relativt ikke-signifikante P-værdier (>0.5; Fig. 10-panel 3). Veje med relativt højt antal metabolitter (n = 5) var: proteinfordøjelse og absorption, b af plante sekundære metabolitter, b af antibiotika og B af aminosyrer. I S4 identificerede vi 58 differentielt regulerede metabolitter, der var forbundet med et stort antal veje, hovedsagelig med EF 1.0 og forholdsvis høje signifikansværdier (for det meste P > 0.5; Fig. 10-panel 4). 3) var: phenylpropanoid b, phenylalanin, tyrosin og tryptophan b, glucosin b, B af alkaloider afledt af shikimatvej og 2-oksocarboksylisk m. i den modne frugt (S5) identificerede vi 39 differentielt regulerede metabolitter forbundet med et stort antal veje, men for det meste med lave P-værdier og kun 1 metabolit pr.vej (Fig. 10-panel 5). Veje med mere end 1 metabolit var: proteinfordøjelse og absorption, phenylpropanoid b, mineralabsorption, centralt kulstof m i kræft, b af sekundære metabolitter, b af phenylpropanoider og aminoacy tRNA b. hovedkomponentanalysen (PCA) af alle data (2 arter, der er 5 stadier, er 5 biologiske replikater) afslørede høj lighed mellem biologiske replikater (klyngedannelse) og bekræftet bemærkelsesværdig variation mellem forskellige frugtmodningstrin for begge arter (Fig. 10-panel 6).

sammenlignende analyse af Kegg metabolisk vej berigelse. De øverste 15 veje beriget i L. ruthenicum i sammenligning med L. barbarum er vist til venstre (rød), og dem beriget i L. barbarum i sammenligning med L. ruthenicum til højre (grøn). Udviklingsstadier (1-5) er angivet i figuren. s-værdien er en FDR-justeret p-værdi.
Metabolome-individuelle metabolitter
listen (supplerende datasæt S6) over de mest differentielt regulerede metabolitter mellem de to arter udviste en vis variation blandt de fem frugtudviklingsfaser. Interessant nok var Fructose 1-phosphat den mest opregulerede metabolit i LR sammenlignet med LB i alle fem faser: log2 Fold ændring = 6,3, 7,6, 7,7, 8,1 og 6,5 (trin 1 til 5 henholdsvis). Med hensyn til metabolitterne stærkt opreguleret i LB var der mere variation mellem faser: i S1 var forskellene ret små med 9-Decenol som den mest opregulerede metabolit (log2 Fold ændring = 2,7; sammenlignet med LR). Analyser af S2 og S3 gav meget kongruente resultater med phenol (henholdsvis 3,7 og 3,2) som den mest opregulerede metabolit. I S4 var indoksylsulfat (4,7) den øverste metabolit opreguleret i LB. Et metabolisk skift blev observeret i den modne frugt (S5), hvor listen over metabolitter opreguleret i LB blev toppet af stearoylcarnitin (7.1), Metoksyeddikesyre (5.3), S-Methyl-5′-thioadenosin (4.7), lisinopril (4.7), adenosin 3′, 5′-cyklisk fosfat (cAMP) (4.7) osv. Andre metabolitter stærkt opreguleret i LR (bortset fra Fructose 1-phosphat) var naringin (6.2), lauroyl-CoA (4.8), L-Phneylalanin (4.6) osv.