a L. barbarum és a L. ruthenicum gyümölcseit öt fejlődési szakaszban gyűjtöttük össze, a fiatal gyümölcsöktől (10 nappal a virágzás után) az érett (érett) gyümölcsökig (34-45 nappal a virágzás után), és tanulmányoztuk azok transzkriptomát és metabolomát.
- RNS-seq de novo az unigenes összeállítása és funkcionális annotációja
- Lycium barbarum (LB): pairwise interstage analysis
- LB Transcriptome
- lb Metabolom
- Lycium ruthenicum (LR): pairwise interstage analysis
- LR Transcriptome
- LR Metabolom
- különböző fejlődési szakaszokban a DEGs interspecifikus összehasonlító elemzése
- összes DEGs a gyümölcsfejlődés során
- transzkriptom-utak
- transzkriptóm-egyedi gének
- Metabolom – utak
- Metabolom-egyedi metabolitok
RNS-seq de novo az unigenes összeállítása és funkcionális annotációja
összesen 30 cDNS-könyvtárat készítettünk a L. barbarum és a L. ruthenicum gyümölcseiből, három biológiai ismétléssel (három gyümölcs három fáról) minden időpontban: 2 faj / 5 időpont / 3 biológiai másolat. A mintákat lb/LR(1-5)-(1-3) jelöléssel látták el, ahol LB az L. barbarum és LR az L. ruthenicum, az 1-5 a gyümölcs fejlődési szakaszai (S1–S5), az 1-3 pedig egyedi minták (biológiai ismétlések); így például az LB1-1 az L. barbarum, 1. mintavételezett fejlődési szakasz (S1), 1.gyümölcsminta (háromból). Több mint 1,72 milliárd páros végű olvasatot generáltunk ehhez a 30 cDNS könyvtárhoz, ami átlagosan 57,2 millió olvasásnak felel meg mintánként (S1 kiegészítő adatkészlet). A szigorú minőségértékelés és adatszűrés összesen 801 766 kiváló minőségű olvasást eredményezett, átlagos hossza 730, N50 pedig 1107 bp (1.táblázat). Végül az átiratokból összesen 326 276 unigént kaptunk, amelyek átlagos hossza 596 bp, az N50 pedig 847 bp volt (1.táblázat). A 30 minta RNS-seq adatainak korrelációs együtthatói az eredmények nagyon jó konzisztenciáját jelzik a biológiai replikációk között (ábra. 2).

az RNS-seq adatok korrelációs együtthatóinak hőtérképe 30 L. barbarum (LB) és L. ruthenicum (LR) gyümölcsből öt különböző fejlődési szakaszban. A mintákat LB/R1–5_1–3 címkével látják el, ahol LB L. barbarum, LR L. ruthenicum, 1-5 a gyümölcs fejlődési szakaszai, 1-3 egyedi minta. A mintákat hierarchikus csoportosítás szerint csoportosítottuk; a hőtérkép felett és bal oldalán található dendrogramok jelzik a minták rokonságát.
a nyilvános adatbázisokkal lekérdezett 326 276 unigén közül összesen 193 021 (59,15%) egyezett géneket és/vagy fehérjéket legalább egy adatbázisban, és 12 171-et (3,73%) jegyeztek fel az összes adatbázisban. A legtöbb unigént (149 863, 45,93%) az NT adatbázisban jegyezték fel, a legkevesebb (24 017; 7,36%) pedig a KOG adatbázisban.
Lycium barbarum (LB): pairwise interstage analysis
LB Transcriptome
a DEGs legnagyobb számát az 1.szakasz összes páronkénti összehasonlításában, valamint a 2. vs. 5. szakasz összehasonlításában azonosították (mind > 10 000 DEGs; 2. táblázat). A legkisebb számokat a 3.vs. 4. és a 4. vs. 5. szakasz összehasonlításaiban azonosították (255-257). A deg-k hőtérkép-elemzése az LB-ben azt mutatja, hogy a korai fejlődési szakaszban (1 + 2) és a későbbi szakaszokban (3-5) meglehetősen eltérő génkészletek voltak erősen szabályozva (ábra. 3A). A minta rokonsági elemzése azt mutatja, hogy a mintákat két kládra lehet osztani (1 + 2 szakasz és 3 + 4 + 5), az utóbbi klád további két kládra oszlik: 3 + 4 és 5.szakasz. Ezeknek a deg-knek az intraspecifikus Kegg funkcionális osztályozási elemzése 15 utat azonosított szignifikánsan (P < 0,05), amelyeket a különböző fejlődési szakaszok különbözőképpen szabályoztak (S2 kiegészítő adatkészlet). Különösen erősen differenciáltan szabályozták a növényi hormon szignál transzdukcióját, a fenilpropanoid bioszintézist (B.), a linolsav metabolizmust (m.), a keményítőt és a szacharózt (m.).’, és ‘zeatin b.’ (ábra. 3A).

a Lycium barbarum (A panel) és az L. ruthenicum (B panel) gyümölcsökben található differenciálisan expresszált gének (DEGs) Hőtérképei és funkcionális útvonalainak elemzése. A hőtérképeket a DEGs (y tengely) és az egyes minták (x tengely) hierarchikus elemzésével állítottuk elő, ahol a hőtérkép felett és bal oldalán található dendrogramok jelzik a minták rokonságát. A mintákat LB/R_1–5_1–3 címkével látják el, ahol a faj rövidítése (LB vagy LR) után következik a gyümölcs fejlődési szakasza (1-5), majd a minta száma (1-3). A két faj mind az öt fejlődési szakaszában a DEGs intraspecifikus Kegg útvonal-elemzését a hőtérképek jobb oldalán mutatjuk be. Csak a top 15 dúsított utak vannak felsorolva. q-érték egy FDR-korrigált p-érték.
az adatok mélyebb elemzéséhez, a legjelentősebb szabályozott utak összehasonlítására összpontosítottunk az egymást követő fejlődési szakaszokban. Az első páronkénti összehasonlításban (S1 vs S2) a’ phenylpropanoid b. ‘volt a leginkább differenciálisan szabályozott út, amelyet a’ keményítő és szacharóz m. ‘ követett (ábra. 4). Nagyon nagy számú DEGs-t (>100) azonosítottak mindkét útvonalon. Hasonló eredményt figyeltek meg a következő páros összehasonlításban, S2 vs.S3, de a DEGs (>80) keményítő és szacharóz m meglehetősen nagy száma ellenére.’valamivel alacsonyabb q-értéket mutatott. Az S3 vs. S4 összehasonlításban a fotoszintetikus organizmusok szénkötése volt a legjelentősebb szabályozott út, de a gének száma sokkal alacsonyabb volt. Az utolsó párban az S4 vs. S5, a ‘zeatin b.’, a flavonoid b.’, a zsírsav b.’ és a ‘galaktóz m.’ voltak a legjelentősebb szabályozott útvonalak, de egyik útvonal sem mutatott kettőnél több DEGs-t.

LB transzkriptom: páronként szakaszközi Kegg metabolikus út dúsítása. A) 1. szakasz vs. 2. szakasz. B) 2. szakasz vs. 3. szakasz. C) 3. szakasz vs. 4. szakasz. D) 4. szakasz vs. 5. szakasz. Top 15 (vagy az összes, Ha összesen <15) jelentősen dúsított utak jelennek meg. A sáv színe a q-érték nagyságát jelöli (FDR-korrigált p-érték). A színdiagram az ábrán látható.
lb Metabolom
páronkénti fejlődési stádiumú összehasonlításokat végeztünk a metabolitok dúsulásának azonosítására az összes stádiumpár között. A differenciálisan szabályozott metabolitok száma az LB fejlődési szakaszainak páros összehasonlításában 66-tól (3.vs. 4. stádium) 129-ig (2. vs. 5. stádium) változott. Az egymást követő szakaszok összehasonlításában a legnagyobb számot az 1.és a 2. szakasz között figyelték meg (117). Ezeket nagyszámú útvonalhoz rendelték; a legtöbb metabolit a másodlagos metabolitok b.-jához (szintén a legalacsonyabb p-értékhez), majd ezt követte a fehérjék emésztése és felszívódása és az aminosavak b. – je (ábra). 5. Kiegészítő Adatok). Az egymást követő szakaszban pár összehasonlítás (S2 vs. S3), az ‘ABC transzporterek’ volt a legjelentősebb dúsított út, amelyet a ‘purin m.’ követett. Az S3 vs. S4 páros összehasonlításban a ‘mikrobiális m. változatos környezetben’ volt a legjelentősebb dúsított út, amelyet a ‘carbapenem m.’követett. Az S4 vs. S5 Párok összehasonlításában viszonylag sok útvonal mutatott hasonló eredményeket (két metabolit és hasonló p-érték), de figyelemre méltó az ‘izoflavonoid b.’ és a ‘flavonoid b.’ megjelenése közöttük (ábra. 5. Kiegészítő Adatok).

LB metabolomikus adatok: páronként szakaszközi Kegg metabolikus út dúsítása. A) 1. szakasz vs. 2. szakasz. B) 2. szakasz vs. 3. szakasz. C) 3. szakasz vs. 4. szakasz. D) 4. szakasz vs. 5. szakasz. A gazdag faktor az útvonalban szignifikánsan szabályozott metabolitok számának és az ezen útvonalon feljegyzett metabolitok teljes számának aránya (tartomány = 0-1,0). A pont mérete a megfelelő útvonalon jelentősen dúsított metabolitok számát, A pont színe pedig a P-értéket jelöli (mindkét legenda látható az ábrán).
ami az egyes metabolitokat illeti, az S1 vs. S2 összehasonlításban az S1-ben szabályozott metabolitok listáját a trehalóz, a Glaktinol és az L-almasav vezette (mind a 20. Az olajsav, a 2-Oxoadipinsav és a sztearinsav voltak a leginkább felszabályozott metabolitok az S2-ben (mind 65-18 log2fc). Az S2-ben az S3-hoz képest leginkább szabályozott metabolitok listáját a dihidroxi-aceton, a Lizopc(18:1(9Z)) és az adenin (mind 16-17, 5 log2fc) vezette. Az S3-ban (az S2-hez képest) a leginkább szabályozott metabolitok listáját a trehalóz, a Galaktinol és az L-almasav vezette (mind a 19-20. Az S3 vs. S4 összehasonlításban az S3-ban a leginkább szabályozott metabolitok listáját az L-almasav, a DL-arginin és az olajsav (mind 68-19 log2fc), míg az S4-ben a leginkább szabályozott metabolitok listáját a D-mannóz, az N-acetil-Laktózamin és a Lizopc(18:1(9Z)) vezette az LB-ben (mind 16 log2fc). Az S4 vs. S5 összehasonlítás, az S4 leginkább szabályozott metabolitjainak listáját a trehalóz, a palmitinsav, az N-acetil-laktóz-amin (mind a 17-18 log2fc) vezette LB-ben. Az S5-ben a leginkább szabályozott metabolitok listáját az L-Norleucin, az antranilsav (L1-Vitamin) és a DL-arginin (mind 16-18 log2fc) (S3 kiegészítő adatkészlet) vezette.
Lycium ruthenicum (LR): pairwise interstage analysis
LR Transcriptome
a DEGs Hőtérkép elemzése az LR-ben azt mutatja, hogy a korai fejlődési szakaszban (1 + 2) és az érett gyümölcsben (5.stádium) szinte teljesen különböző génkészleteket szabályoztak erősen, a második szakasz után látszólagos transzkriptomikus reset következett be (ábra. 3B). A minta rokonsági elemzése azt mutatja, hogy a mintákat két kládra lehet osztani (1 + 2 szakasz és 3 + 4 + 5), az utóbbi klád további két kládra oszlik: 3 + 4 és 5.szakasz. Ezeknek a deg-knek az intraspecifikus Kegg funkcionális osztályozási elemzése 35 utat azonosított szignifikánsan (P < 0,05), amelyeket a különböző fejlődési szakaszok különbözőképpen szabályoztak (S2 kiegészítő adatkészlet). Különösen erősen differenciáltan szabályozták a ‘fotoszintézist’, ‘linolsav m.’, ‘izokinolin alkaloid b.’, ‘flavonoid b.’ és ‘keményítő és szacharóz m’ (ábra. 3B). A DEGs-ek legnagyobb számát az 1. szakasz legtöbb páronkénti összehasonlításában azonosították (1 vs.2 kivétel volt), valamint a 2. vs. 5 összehasonlításban (mind >5000 DEGs; 2. táblázat). Messze a legkisebb számot azonosították az S3 vs. S4 összehasonlításban (39). Az S1 vs S2 páros összehasonlításban az ‘amino cukor és nukleotid cukor m.’ volt a leginkább differenciálisan szabályozott út, amelyet a ‘linolsav m.’ követett (ábra. 6). Az S2 vs. S3 páros összehasonlításban az izokinolin alkaloid b., a tirozin m., a fenilpropanoid B. és a flavonoid B. Mindkét páros összehasonlításban a DEGs-ek legnagyobb számát (>15, illetve >35) a keményítőben, illetve a szacharóz m-ben azonosították. Az S3 vs. S4 összehasonlítás, ‘(alfa -) linolsav m. ‘ volt a legjelentősebb szabályozott út, de a gének száma sokkal alacsonyabb volt. Az S4 vs. S5 összehasonlításban a ‘fenilpropanoid b’ (szintén a legtöbb DEGs) és a ‘linolsav m.’ voltak a legjelentősebb szabályozott útvonalak.
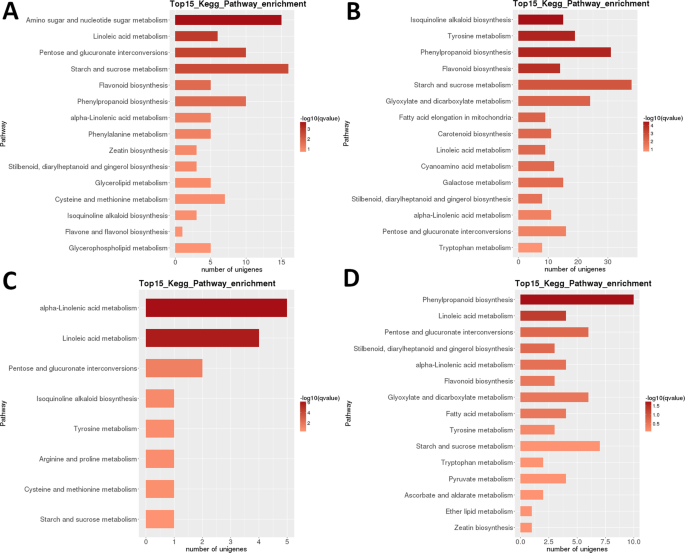
LR transzkriptom: páronként szakaszközi Kegg metabolikus út dúsítása. A) 1. szakasz vs. 2. szakasz. B) 2. szakasz vs. 3. szakasz. C) 3. szakasz vs. 4. szakasz. D) 4. szakasz vs. 5. szakasz. Top 15 (vagy az összes, Ha összesen <15) jelentősen dúsított utak jelennek meg. A sáv színe a q-érték nagyságát jelöli (FDR-korrigált p-érték, az ábrán látható színdiagram).
LR Metabolom
a differenciálisan szabályozott metabolitok száma az LR fejlődési szakaszainak páronkénti összehasonlításában 66-tól (3.vs. 4. stádium) 133-ig (2. vs. 5. stádium) változott (S3 kiegészítő adatkészlet). Az egymást követő szakaszok összehasonlításában a legnagyobb számot az 1.és a 2. szakasz között figyelték meg (117). Ezeket nagyszámú útvonalhoz rendelték; a legtöbb metabolit a szekunder metabolitok b-jéhez van hozzárendelve, ezt követi a fehérjék emésztése és felszívódása, az aminosavak b-je és a flavonoid b-je. Az S2 vs. S3 összehasonlításban az ‘ABC transzporterek ‘volt a legjelentősebb dúsított útvonal, amelyet a’purin m’ követett. Az S3 vs. S4 páros összehasonlításban a ‘mikrobiális m. változatos környezetben’ volt a legjelentősebb dúsított út, amelyet a ‘carbapenem m.’követett. Az S4 vs. S5 Párok összehasonlítása, viszonylag sok útvonal mutatott hasonló eredményeket (2 metabolit és hasonló p-értékek), de figyelemre méltó az ‘izoflavonoid b.’ és a ‘flavonoid b.’ megjelenése közöttük (ábra. 7. Kiegészítő Adatok).

LR metabolomikus adatok: páronként szakaszközi Kegg metabolikus út dúsítása. A) 1. szakasz vs. 2. szakasz. B) 2. szakasz vs. 3. szakasz. C) 3. szakasz vs. 4. szakasz. D) 4. szakasz vs. 5. szakasz. A gazdag faktor az útvonalban szignifikánsan szabályozott metabolitok számának és az ezen útvonalon feljegyzett metabolitok teljes számának aránya (tartomány = 0-1,0). A pont mérete a megfelelő útvonalon jelentősen dúsított metabolitok számát, A pont színe pedig a P-értéket jelöli (mindkét legenda látható az ábrán).
az egyes metabolitok (S3 kiegészítő adatkészlet) tekintetében az S1 vs. S2 összehasonlítás, Az S1-ben szabályozott metabolitok listáját a trehalóz, a Galaktinol és az L-almasav (69-21 log2FC) vezette, míg az S2-ben az olajsav, a 2-Oxoadipinsav és a sztearinsav voltak a leginkább szabályozott metabolitok (15-18 log2fc). Az S2 vs. S3 összehasonlításban a dihidroxi-aceton, az Indoxil-szulfát és az N-acetil-Laktózamin voltak a leginkább felszabályozott metabolitok az S2-ben (6-17-19, 5 log2fc), valamint a trehalóz, Galaktinol és L-almasav (19-21 log2fc) az S3-ban. Az S3 vs. S4 összehasonlítás, az L-almasavat, a DL-arginint és az olajsavat az S3-ban (6-16-19 log2FC), az 1,7-dimetil-xantint, a D-mannózt és az N-acetil-laktózt (15-17 log2fc) az S4-ben szabályozták. Az S4 vs. S5 összehasonlításban a Flavin mononukleotidot, a trehalózt és az Izoferulinsavat az S4-ben (6-18-20 log2fc) szabályozták, míg a PG(16:0/18:1(9Z)), a D-prolin és a DL-arginin volt a leginkább szabályozva (mind 16-18 log2fc) metabolitok.
különböző fejlődési szakaszokban a DEGs interspecifikus összehasonlító elemzése
összes DEGs a gyümölcsfejlődés során
interspecifikus páros szakasz összehasonlítás (LR1 vs.LB1, LR2 vs. LB2 stb.) azt mutatja, hogy 928 DEGs osztoztak mind az öt pár (ábra. 8A). A legtöbb DEGs-t a 3.szakaszban (3989), a legkevesebbet a 4. szakaszban (2825) azonosították (ábra. 8B); mivel az egy párra jellemző DEGs-ek legnagyobb számát a 3. (574.), az 1. és az 5. (mindkettő 554.) szakaszban, a legalacsonyabbat pedig a 4. szakaszban (126. ábra) figyelték meg (ábra. 8A). A felfelé és lefelé szabályozott deg – ek száma viszonylag hasonló volt az egyes páros szakaszok összehasonlításában; például az 5. szakaszban az LR-ben 1668 deg-et szabályoztak, az LB-hez képest pedig 1670 deg-et szabályoztak. 8B). A másik négy szakaszban azonban a felszabályozott gének száma kissé (93-189 DEGs) magasabb volt.

az L. barbarum (LB) és az L. ruthenicum (LR) gyümölcsei között differenciáltan expresszált gének (DEGs). (A) részletes szakaszonként (1-5) összehasonlítások (LB vs.LR). (B) a szabályozatlan (piros) és szabályozatlan (zöld) deg-k száma az LR-ben az LB-hez képest öt vizsgált fejlődési szakaszban.
transzkriptom-utak
a DEGs Hőtérkép-elemzése azt mutatja, hogy két faj gyümölcse nagyon eltérő génexpressziós profilt mutat minden fejlődési szakaszban, de a biológiai replikációk nagyon hasonló profilokat mutattak, ami korlátozott mennyiségű egyedi variabilitást jelez az egyes fejlődési szakaszokban (kiegészítő ábrák: ábra. S9). A KEGG útvonal dúsulásának összehasonlító elemzése azt mutatja, hogy csak néhány útvonal volt következetesen erősen dúsítva (a génszabályozás szempontjából) az LR-ben az LB-hez képest mind az öt fejlődési szakaszban (ábra. 9). Különösen a növényi hormon jelátvitel (2.-legmagasabb Az S1-ben, 8.-legmagasabb az S2-ben, a legmagasabb az S3-ban, S4-ben és S5-ben) és a növényi kórokozó kölcsönhatás (a legmagasabb Az S1-ben, 3. – legmagasabb az S2-ben, 2. – legmagasabb az S3-ban, 4. – legmagasabb az S4-ben, és 15. – legmagasabb az S5-ben) viszonylag erősen szabályozottak voltak minden szakaszban. A fenilpropanoid bioszintézist (nem a top 15-ben az S1-ben, a legmagasabb az S2-ben, a 3.-a legmagasabb az S3-ban, a 7.-a legmagasabb az S4-ben, a 11.-a legmagasabb az S5-ben), az ubiquinont és más terpenoid-kinon bioszintézist (nem a top 15-ben az S1-ben, a 6. – a legmagasabb az S2-ben, a 7. – a legmagasabb az S3-ban, a 2. – a legmagasabb az S4-ben, a 6. – a legmagasabb az S5-ben) az első kivételével minden szakaszban viszonylag erősen szabályozták. A Flavonoid bioszintézis útvonala nem volt nagymértékben dúsítva a korai szakaszban (nem a top 15-ben az S1-ben, a 14. – legmagasabb az S2-ben), és a késői szakaszokban (3. -4. – legmagasabb a 3-5. szakaszban). az (alfa-)linolsav metabolizmus erősen dúsított volt a középső szakaszokban (9. – legmagasabb Az S1-ben, 2.-legmagasabb az S2-ben, 5. és 6. – legmagasabb az S3-ban, 8. – legmagasabb az S4-ben, nem az első 15-ben az S5-ben).

a Kegg metabolikus út dúsulásának összehasonlító elemzése. A L. ruthenicumban dúsított felső 15 útvonal a L. barbarumhoz képest a bal oldalon (piros), a L. barbarumban dúsított utak a L. ruthenicumhoz képest a jobb oldalon (zöld). A fejlődési szakaszokat (1-5) az ábra mutatja. q-érték egy FDR-korrigált p-érték.
között az utak downregulated L. ruthenicum képest L. barbarum (ábra. 9) jelentős változásokat figyeltek meg a korai (1.és 2.) fázisok között, amikor a cianoaminosav-metabolizmus és a karotinoid-bioszintézis a legnagyobb mértékben csökkent, és a késői (4. és 5.) fázisok között, amikor a hólyagos transzport, a nikotinát-és nikotinamid-metabolizmus, valamint a porfirin-és klorofill-metabolizmus pergő kölcsönhatásai következetesen viszonylag erősen csökkentek voltak.
transzkriptóm-egyedi gének
a leginkább differenciálisan expresszált gének közül néhány fejlődési szakasz-specifikus volt (azaz csak a korai vagy a késői fejlődési szakaszban szabályozták erősen differenciálisan), de néhányat következetesen erősen szabályoztak mind az öt vizsgált szakaszban (S4 kiegészítő adatkészlet). Számos immunitással kapcsolatos gén nagyon erősen szabályozva van az LR-ben, összehasonlítva az LB-vel a korai fejlődési szakaszban, a fejlődési szakasz-specifikus expressziós minta példái közé tartozik: a kitináz volt a 2. legmagasabb szabályozott fok Az S1-ben (13,43-szoros), a legmagasabb az S2-ben (13,89-szeres), de a későbbi szakaszokban nem volt fok. Hasonlóképpen, az EIX receptor 1/2 szintén nagyon erősen szabályozva volt a korai szakaszban, a legmagasabb Az S1 – ben (13,70) és a 2.legmagasabb az S2-ben (10,90), de a későbbi szakaszokban sem azonosították DEG-ként. Néhány növekedéssel kapcsolatos gén is hasonló expressziós mintázatot mutatott: a foszfoglicerát-kináz (PGK) az első három szakaszban (13,14, 12,87 és 12) a maroknyi leginkább felszabályozott gén között volt.77), de a későbbi szakaszokban szintén nem azonosították DEG-ként. A CCR4-NOT transzkripciós komplex 7/8 alegység (CNOT7 / 8) szintén nagyon hasonló expressziós mintát mutatott: az első három szakaszban erősen szabályozva, a 4.és az 5. szakaszban pedig nem DEG. Számos flavonoid és fenilpropanoid bioszintézishez kapcsolódó gén fordított fejlődési szakasz-specifikus expressziós mintázatot mutatott, viszonylag alacsony expresszióval a korai szakaszban, és nagyon magas a későbbi szakaszokban. Példák: bifunkcionális dihidroflavonol 4-reduktáz/flavanon 4-reduktáz (DFR), amelyet az LR-ben kissé szabályoztak az S1-ben (2.25), nem DEG az S2-ben, erősen felszabályozott az S3-ban (7,79), és a 3.legmagasabb felszabályozott gén az S4-ben (14,25) és az S5-ben (16,03). Ennek a génnek a paralógja szinte azonos mintázatot mutatott: kissé felfelé szabályozva az S1-ben (2,44), nem egy fok az S2-ben, erősen szabályozva az S3-ban (7,40), a 6.legmagasabb felfelé szabályozva fok az S4-ben (13,26), az 5. pedig az S5-ben (14,59). Hasonlóképpen, a flavonoid 3′, 5 ‘- hidroxiláz (F3 ‘ 5 ‘ H) nem volt DEG az első két szakaszban, erősen szabályozva az S3-ban (6,69), az 5.legmagasabb szabályozású gén az S4-ben (13,42), a 4. legmagasabb pedig az S5-ben (15,05). A Flavonoid O-metiltranszferáz (OMT) nem volt DEG az S1-ben, de az S2-ben már közepesen magas upregulációs szintet mutatott (4,32), az S3-ra már a harmadik legmagasabb upregulált deg (13,30), és a legmagasabb upregulált gén volt az S4-ben (18,73) és az S5-ben (18,10). A leukoantocianidin-dioxigenáz (LDOX; antocianin bioszintézis) nem volt DEG az S1-ben és az S2-ben, amelyet a későbbi szakaszokban magas vagy nagyon magas upreguláció követett (sorrendben 5,63, 9,44, 11,56). Két kalkon-szintáz paralógot (CHS és CHS2; flavonoid bioszintézis) szintén nem szabályoztak erősen Az S1 és S2 (CHS2: nem a DEG, CHS: -1,14 az S1-ben, nem egy fok az S2–ben), de az S3-S5-ben mindkét gén közepes-magas vagy magas upregulációt mutatott (CHS2: 5,32, 7,84, 6,00; és CHS: 4,67, 7,01, 6,82; illetőleg). Ezeket a géneket választottuk ki a qPCR analízishez, és az eredmények nagymértékben egybevágnak az RNS-seq adatokkal (kiegészítő eredmények; S5 kiegészítő adatkészlet). Végül a citokinin-dehidrogenáz, a zeatin bioszintézishez kapcsolódó gén szintén egyre inkább szabályozódott az utolsó három szakaszban (2,6–5,7).
néhány gén azonban következetesen differenciálisan expresszálódott mind az öt vizsgált szakaszban. A példák között szerepelt néhány immunitással kapcsolatos gén is, például a glutation S-transzferáz két paralógja, amelyek az LR-ben erősen szabályozottak az LB-hez képest minden szakaszban: 9,38 és 8,58 (az összes érték a megfelelő sorrendben történő hajtásváltozásként jelenik meg) az S1-ben, 6,30 és 6,34 az S2-ben, a 2.és 7. legmagasabb szabályozottsági DEGs az S3-ban (14,08 és 12,70), a 2. és 4. legmagasabb az S4-ben (15,71 és 14.16) és a 2. és 6. legmagasabb az S5-ben (16.48 és 14.40). Az rpm1 növénybetegségekkel szembeni rezisztencia fehérje mind az öt szakaszban erősen szabályozva volt (S1 = 13,15; S2 = 12,08; S3 = 13,11, S4 = 12,81; S5 = 13:94). Az összes fejlődési szakaszban következetesen differenciálisan expresszált gének között voltak olyanok is, amelyek az aminosav-anyagcseréhez kapcsolódtak, de mintázatuk megfordult a korábbi példákhoz képest: az LR-ben az LB-hez képest magas downregulációt mutattak. Ilyen például az acetil-CoA Acil-transzferáz 1 (aat1; valin, leucin és izoleucin degradáció), amelynek időbeli profilja egyre nagyobb mértékű downregulációval jár, kezdve -7,0−től Az S1-ben < – 10-szeresig az utolsó három szakaszban. Az arginin és prolin metabolizmushoz társuló prolin iminopeptidáz az LR-ben minden szakaszban erősen csökkent volt: S1 = -9,75, S2 = -10,89 (3.legmagasabb), S3 = -11,05 (4. legmagasabb), S4 = -10,01 és S5 = -11,98 (3. legmagasabb). Végül az 5–metil-tetrahidropteroil-triglutamát-homocisztein-metiltranszferáz (metE) következetesen rendkívül erősen csökkent az LR-ben minden szakaszban: a 2.legmagasabb Az S1-ben (-11,76), a legmagasabb az S2-ben (-11,74), a 3. legmagasabb az S3-ban (-11,43), a legmagasabb az S4-ben (-12,36) és a 2. legmagasabb az S5-ben (-12,83). Két DNS-replikáció és transzkripció-asszociált gén is erősen csökkent az LR-ben minden szakaszban: a GTP-kötő nukleáris fehérje Ran (RAN; -10,0 -12,0) és a replikációs faktor A1 (RFA1) (-8,0-12,0). Néhány növekedéssel és stresszel kapcsolatos gén szintén következetesen erősen csökkent az LR-ben: heterogén nukleáris ribonukleoprotein A1/A3 (hnRNP; -7-11) és hősokk 70 kDa fehérje 1/8 (HSPA1_8) S1 = -4,95, S2 = -8,88, S3 = -11,48 (2.legmagasabb), S4 = -9,36, S5 = -12,89 (a legmagasabb). Érdekes módon egy fenilpropanoid bioszintézishez kapcsolódó gén, a sikimát-hidroxi-cinnamoil-transzferáz (HCT) szintén következetesen erősen csökkent az LR-ben: S1 = -6,82, S2 = -8,14, S3 = -11,71 (a legmagasabb), S4 = -11,00 (3.legmagasabb), S5 = -11,91 (4. legmagasabb). Az antocianin bioszintézis kulcsfontosságú szabályozója, a myb114 transzkripciós faktor azonban mind az öt fejlődési szakaszban erősen szabályozódott az LR-ben: 6,11, 4,69, 7,47, 9,05, illetve 8,95 (S1–S5).
Metabolom – utak
a metabolikus utak összehasonlító interspecifikus szakaszonkénti elemzését is elvégeztük (ábra. 10). Az első fejlődési szakaszban (S1) 39 differenciálisan szabályozott metabolitot azonosítottunk. A top 20 útvonal közül ezek a metabolitok társultak, több közülük aminosavakkal társult, de a metabolitok teljes száma útvonalonként viszonylag kicsi volt (1-2), és a P-értékek nem utaltak nagy jelentőségre (ábra. 10-panel 1). Különösen a B6 – vitamin metabolizmus (m) és a mikrobiális m különböző környezetekben viszonylag magas P-értékeket, 1,0-es dúsítási faktort (EF) és 2 azonosított metabolitot mutatott. Az S2-ben 58 differenciálisan szabályozott metabolitot azonosítottunk, amelyek csak négy útvonalhoz kapcsolódnak: triptofán m, fenilpropanoid bioszintézis (B), fenilpropanoidok b (Ezek két különböző útvonal a KEGG adatbázisban), valamint fenilalanin, tirozin és triptofán b (minden EF = 1,0, 2-3 metabolit és p < 0,5; ábra. 10-panel 2). Az S3-ban 59 differenciálisan szabályozott metabolitot azonosítottunk, amelyek 19 útvonalhoz kapcsolódnak, leginkább az EF 1.0-val, de viszonylag nem szignifikáns P-értékekkel (>0.5; ábra. 10-panel 3). A viszonylag nagy számú metabolitot (n = 5) tartalmazó útvonalak a következők voltak: fehérje emésztés és felszívódás, növényi másodlagos metabolitok b-je, antibiotikumok b-je és aminosavak b-je. Az S4-ben 58 differenciálisan szabályozott metabolitot azonosítottunk, amelyek nagyszámú útvonalhoz kapcsolódnak, többnyire az EF 1.0-val, és viszonylag nagy szignifikanciájú értékeket (többnyire P > 0.5; ábra. 10-panel 4). A viszonylag nagy számú metabolitot (n 6) tartalmazó útvonalak a következők voltak: fenilpropanoid b, fenilalanin, tirozin és triptofán b, glükozinát B, sikimát útvonalból származó alkaloidok b és 2-oxokarboxil m. az érett gyümölcsben (S5) 39 differenciálisan szabályozott metabolitot azonosítottunk, amelyek nagyszámú útvonallal társultak, de többnyire alacsony P-értékekkel és útvonalonként csak 1 metabolittal (ábra. 10-panel 5). Az 1-nél több metabolitot tartalmazó utak a következők voltak: fehérje emésztés és felszívódás, fenilpropanoid b, ásványi abszorpció, központi szén m a rák, B másodlagos metabolitok, B fenilpropanoidok, és aminoacy tRNS b. a főkomponens analízis (PCA) az összes adat (2 faj ~ 5 szakasz ~ ~ 5 biológiai ismétlődések) kiderült, nagy hasonlóság között biológiai ismétlődések (klaszterezés), és megerősítette jelentős variabilitás a különböző gyümölcs érési szakaszában mindkét faj (ábra. 10-panel 6).

a Kegg metabolikus út dúsulásának összehasonlító elemzése. A L. ruthenicumban dúsított felső 15 útvonal a L. barbarumhoz képest a bal oldalon (piros), a L. barbarumban dúsított utak a L. ruthenicumhoz képest a jobb oldalon (zöld). A fejlődési szakaszokat (1-5) az ábra mutatja. q-érték egy FDR-korrigált p-érték.
Metabolom-egyedi metabolitok
a két faj között a leginkább differenciálisan szabályozott metabolitok listája (S6 kiegészítő adatkészlet) némi eltérést mutatott az öt gyümölcsfejlődési szakasz között. Érdekes módon a fruktóz-1-foszfát volt a legszabályozottabb metabolit az LR-ben, az LB-hez képest, mind az öt szakaszban: log2-szeres változás = 6,3, 7,6, 7,7, 8,1 és 6,5 (sorrendben 1-5.szakasz). Ami az LB-ben erősen szabályozott metabolitokat illeti, több eltérés volt a szakaszok között: az S1-ben a különbségek meglehetősen csekélyek voltak, a 9-Decenol volt a leginkább szabályozott metabolit (log2-szeres változás = 2,7; az LR-hez képest). Az S2 és az S3 analízise erősen egybevágó eredményeket hozott, a fenol (3,7, illetve 3,2) volt a leginkább felszabályozott metabolit. Az S4-ben az Indoxil-szulfát (4.7) volt a legfontosabb metabolit, amelyet az LB-ben szabályoztak. Metabolikus eltolódást figyeltek meg az érett gyümölcsben (S5), ahol az LB-ben szabályozott metabolitok listáját a sztearoilkarnitin (7.1), A Metoxi-ecetsav (5.3), az S-metil-5′ – tioadenozin (4.7), a lizinopril (4.7), adenozin-3′, 5 ‘ -ciklikus foszfát (cAMP) (4.7.) stb. Az LR-ben erősen szabályozott egyéb metabolitok (a fruktóz-1-foszfát kivételével) a naringin (6.2), A lauroil-CoA (4.8), az L-Phneylalanine (4.6) stb.