図4:目の構造。この画像はChirstie(ref. 4)と与えられたソースに従って著作権制限されています(すなわち、それは著者自身の作品ではありません)。
目の構造:光と色を検出する目の中の領域(図4を参照)は、網膜と呼ばれます。 検出の細胞の現在の、棒および円錐形、プロセス情報のcomingthroughレンズのtwotypesは頭脳に視神経の下でそれを送ります。 ロッド細胞(約100万人)は眼に入る明度を検出し、その感度は細胞内で生成されるロドプシンの量に依存する。 但し、ロドプシンはライトへの露出のbybleaching破壊され、従って棒の細胞は低照度のasatの高い照明でだけこの感光性顔料の減らされたレベルaveryの低い感受性 円錐形の細胞は(およそ3百万がある)また光源レベルに敏感ですが、顔料のヨードプシンの使用によって高い照明まで機能を維持します。色の検出は、網膜内に存在する3つのタイプの錐体細胞の機能である:それらの間にそれらは可視スペクトルをカバーする。 これは、eachtypeが赤(長い)、緑(中)、または青(短い)に対応する最大の異なる波長範囲に敏感であるためです。
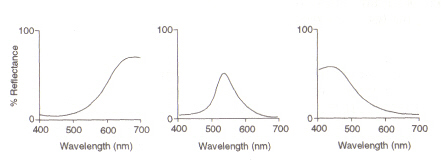
図5:赤、緑、青の錐体細胞の最大値(左から)。 この画像はChirstie(ref. 4)および与えられたソースに従って制限された著作権です(すなわち、それは著者自身の作品ではありません)。
Rodcells内のロドプシンのメカニズム:蛋白質のロドプシンは自然に7つのtrans膜a helicalreceptorsによって形作られる膜間ポケットにあるprotonatedretinal-Schiffの基盤の複合体を含んでいます。 光検出時にロドプシン(11-シス)から全trans網膜への光異性体変化を受けるarod細胞の外側セグメント内にロドプシンの多くの平らなディスクがあります。 5つの短鎖インターメディエート(フローチャート1)を介して起こる光異性化カスケードの後、trans retinalは拡散し、11-cisretinalに変換されてサイクルに再突入する。 このプロセスは暗闇の酸化/異性化に先行している減少のtoall transのレチノールによって起こります。 光励起ロドプシン(4番目の5つの中間体)は、GMPの加水分解に関与する酵素カスケードプロセスを引き起こす。 これは今度は暗闇のNa+の流入に自然に開いている棒の細胞膜の陽イオン特定のchannelswithinを閉め、hyperpolarisationの効果が原因で、内部のsynatic bodysendsは網膜の他のニューロンに神経信号 最後に、カルシウムレベルの光誘導低下は、励起されたニューロンの受動的な”暗い”状態への回復を助け、光の検出時にサイクルが再び開始される。 錐体細胞の光受容体は、発色団として11-cis-網膜を有する七つのa-helicalreceptorsでもある。 検出範囲は緑から赤に変化し,網膜付近の三つの非極性ヒドロキシル含有残基が極性残基に順次置換される。

左図6:ロドプシン内の11-cis網膜の位置。 この画像はStryer(ref. 6)および与えられたソースに従って制限された著作権です(すなわち、それは著者自身の作品ではありません)。
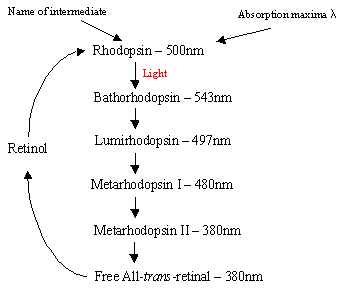
右-Flowchart1: ロドプシンサイクルの5つの中間体。
ロドプシンはどこから来たのですか?
人間はホドプシンを作ることができず、代わりに食品中に見出されるb-カロチンを合成するために外部ソースを使用しています:
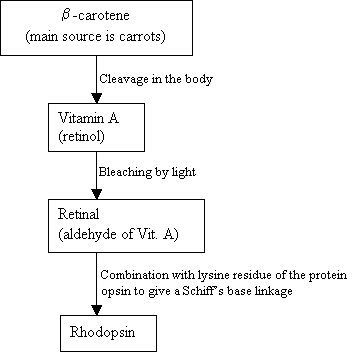 |
フローチャート2:ロドプシン の合成新しい画像=図7:b-カロテン(上)、VitaminA(中)、11-cisレチナール(下)の構造。 |
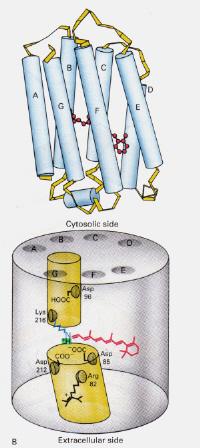
ハロバクテリアに見られるバクテリオロドプシンのメカニズムは、人間の目のロド細胞に見られるロドプシンのメカニズムとどのように異な:今回は、プロトン化された網膜シフの塩基複合体が、そうでなければ二つの隣接するチャンバーによって形成された膜を通ってチャネルを自然にブロ プロトン化されたtrans複合体はAsp-85にプロトンを供与し、同じプロトンを細胞外側に放出することができる。 13-cis構造への光異性化は、シッフの塩基がサイトゾル側のAsp-96残基からプロトンを拾うことを可能にする。 Transへのシス形の再配向時に、異性化とプロトンポンプのサイクルが続く。