l.barbarumおよびL.ruthenicumの果実を、若い果実(開花後10日以上)から成熟(熟した)果実(開花後34-45日)までの5つの発達段階で収集し、それらの果実を研究した。トランスクリプトームとメタボローム。
RNA-seq de novo assembly and functional annotation of unigenes
L.barbarumとL.ruthenicumの果実から合計30のcDNAライブラリーを調製し、各時点で2種×5時点×3の生物学的複製を用いた。 試料は、LB/LR(1−5)−(1−3)と標識され、ここで、LBは、L.barbarumであり、LRは、L.ruthenicumであり、1−5は、果実の発生段階(S1−S5)であり、1−3は、個々の試料(生物学的複製物)であ barbarum、1stサンプリングされた発達段階(S1)、果実サンプルNo.1(三つのうち)。 我々は、サンプル(補足データセットS1)あたり57.2百万読み取りの平均に対応する、これらの30cDNAライブラリのための1.72億ペアエンド読み取りを生成し 厳格な品質評価とデータフィルタリングにより、合計801,766の高品質読み取りが行われ、平均長は730、N50は1107bpでした(表1)。 最後に、596bpの平均長および847bpのN50を有する合計326,276ユニ遺伝子を転写物から得た(表1)。 30サンプルのRNA-seqデータの相関係数は、生物学的複製物間の結果の非常に良好な一貫性を示している(図10B)。 2).

L.barbarum(LB)とL.ruthenicum(LR)果実の30サンプルのRNA-seqデータの相関係数のヒートマップ五つの異なる発達段階で。 試料はLB/R1−5_1−3と標識され、ここで、LBはL.barbarumであり、LRはLである。 ruthenicum、1-5は果実の発達段階であり、1-3個の個々のサンプルである。 サンプルは階層的なクラスタリングによってグループ化され、ヒートマップの上と左の樹枝図はサンプルの関連性を示しています。
公開データベースに対して照会されたすべての326,276unigenesのうち、少なくとも一つのデータベース内の193,021(59.15%)の一致した遺伝子および/またはタンパク質の合計、および12,171(3.73%)は、すべてのデータベースで注釈が付けられた。 UNIGENESの最大数(149,863、45.93%)はNTデータベースで注釈され、KOGデータベースで最も低い数(24,017、7.36%)であった。
Lycium barbarum(LB):ペアワイズ段階間解析
LBトランスクリプトーム
第1段階のすべてのペアワイズ比較、および第2段階と第5段階の比較(すべて>10,000度;表2)で最 最小の数字は、第3対第4および第4対第5段階の比較(255-257)で同定された。 LB中のDegのヒートマップ分析は、かなり異なる遺伝子セットが、初期の発達段階(1+2)および後期段階(3〜5)において高度に上方制御されたことを示す(図 3A)。 サンプル関連性分析は、サンプルが2つのクレード(ステージ1+2およびステージ2)に分けることができることを示しています。3 + 4 + 5)、後者のクレードはさらに二つのクレードに細分された:ステージ3+4とステージ5。 これらのDegの種内KEGG機能分類分析は、15の経路を有意に同定した(P<0.05)差動異なる発達段階(補足データセットS2)の間で調節された。 特に”植物ホルモンシグナル伝達”,”フェニルプロパノイド生合成(b.)”,”リノール酸代謝(m.)”,”でんぷんおよびしょ糖m”が強く調節された。および「zeatin b」(図1)。 3A)。

Lycium barbarum(パネルA)およびL.ruthenicum(パネルB)果実における差動発現遺伝子(Deg)のヒートマップおよび機能経路解析。 ヒートマップは、度(y軸)と個々のサンプル(x軸)の階層分析によって生成され、ヒートマップの上と左のデンドログラムはサンプルの関連性を示します。 サンプルはLB/R_1–5_1–3と表示され、種の頭字語(LBまたはLR)の後に果実の発達段階(1-5)、およびサンプル番号(1-3)が続きます。 二つの種のすべての発生段階におけるDegの種内KEGG経路解析をヒートマップの右側に示した。 上位15の経路のみがリストされています。 q値は、FDR調整されたp値です。
データのより詳細な分析のために、我々は、連続した発達段階で最も有意に調節された経路の比較に焦点を当てた。 最初のペアワイズ比較(S1対S2)では、’フェニルプロパノイドb.’が最も高度に特異的に調節された経路であり、続いて’デンプンおよびショ糖m.’(図10A)が続いた。 4). 非常に多数のDeg(>100)が両方の経路で同定された。 同様の結果は、以下のペアワイズ比較、S2対S3で観察されたが、Deg(>80)’デンプンおよびショ糖mのかなり多数にもかかわらず。’はやや低いq値を示した。 S3とS4の比較では、”光合成生物における炭素固定”が最も有意に調節された経路であったが、遺伝子の数ははるかに低かった。 最後のペアでは、S4対S5、’ゼアチンb.’、フラボノイドb.’、脂肪酸b.’と’ガラクトースm.’最も有意に調節された経路であったが、経路のどれも二つ以上のDegを示

LBトランスクリプトーム:ペアワイズ段階間KEGG代謝経路濃縮。 (A)ステージ1対ステージ2。 (イ)第2ステージ対 ステージ3。 (C)ステージ3対ステージ4。 (D)ステージ4対ステージ5。 上位15(または合計<15の場合はすべて)有意に濃縮された経路が示されている。 バーの色は、q値(FDR調整されたp値)の大きさを表します。 カラーチャートは図に示されています。
LBメタボローム
我々は、すべての段階のペア間の代謝産物の濃縮を識別するためにペアワイズ発達段階の比較を実施しました。 LBの発達段階のペアワイズ比較における差動調節代謝産物の数は66(ステージ3対4)から129(ステージ2対5)に変化した。 連続した段階の比較の最も大きい数は第1および第2段階(117)の間で観察されました。 これらは多数の経路に割り当てられ、”二次代謝産物のb.”(最も低いp値)に割り当てられた代謝産物の数が最も多く、続いて”タンパク質消化吸収”および”アミノ酸のb.”(図。 5;補足の図)。 連続ステージペアの比較では、S2対 S3)、”ABCトランスポーター”は、”プリンm”に続いて、最も有意に濃縮された経路であった。 S3対S4ペアの比較では、”多様な環境における微生物m.”が最も有意に濃縮された経路であり、続いて”カルバペネムm.”が続いた。 S4対S5対の比較では、比較的多数の経路が同様の結果(2つの代謝産物および類似のp値)を示したが、注目すべきは、それらの中で「イソフラボノイド 5;補足の図)。

LBメタボロムデータ: ペアワイズ-ステージ間KEGG代謝経路濃縮。 (A)ステージ1対ステージ2。 (B)ステージ2対ステージ3。 (C)ステージ3対ステージ4。 (D)ステージ4対ステージ5。 リッチ因子は、経路における有意に調節された代謝産物の数と、その経路において注釈された代謝産物の総数との比である(範囲=0から1.0)。 点のサイズは、対応する経路における有意に濃縮された代謝産物の数を表し、点の色はP値を表す(図に示す両方の凡例)。
個々の代謝産物に関しては、S1対S2の比較では、S1でアップレギュレートされた代謝産物のリストは、トレハロース、グラチノールおよびL-リンゴ酸(すべて≥20log2fc)によって突破された。 オレイン酸、2-オキソアジピン酸およびステアリン酸は、S2(すべて≥15から18log2fc)で最も高度にアップレギュレートされた代謝産物であった。 S3と比較してS2で最も高度にアップレギュレートされた代謝産物のリストは、ジヒドロキシアセトン、リゾpc(18:1(9Z))、およびアデニン(すべて≥16〜17.5log2fc) S3の最も高度にアップレギュレートされた代謝産物のリスト(S2と比較して)は、トレハロース、ガラクチノールおよびL-リンゴ酸(すべて≥19から20log2fc)によっ S3対S4の比較では、S3で最も高度にアップレギュレートされた代謝産物のリストは、L-リンゴ酸、DL-アルギニンとオレイン酸(すべて≥18から19log2fc)によっ のS4対… S5の比較では、S4の最も高度にアップレギュレートされた代謝産物のリストは、LBのトレハロース、パルミチン酸、N-アセチルラクトサミン(すべて≥17から18log2fc)によって突破された。 S5で最も高度にアップレギュレートされた代謝産物のリストは、L-ノルロイシン、アントラニル酸(ビタミンL1)およびDL-アルギニン(すべて≥16から18log2fc)(補足データセットS3)によって突破された。
Lycium ruthenicum(LR): ペアワイズ段階間解析
LRトランスクリプトーム
LRのDegのヒートマップ解析は、遺伝子のほぼ完全に異なるセットが高度に発達初期(1+2)と熟した果実(ステージ5)でアップレギュレートされ、明らかなトランスクリプトミックリセットが第二段階の後に発生したことを示している(図。 3B)。 サンプル関連性分析は、サンプルが2つのクレード(ステージ1+2およびステージ2)に分けることができることを示しています。3 + 4 + 5)、後者のクレードはさらに二つのクレードに細分された:ステージ3+4とステージ5。 これらのDegの種内KEGG機能分類分析は、35の経路が有意に同定された(P<0.05)差動異なる発達段階(補足データセットS2)の間で調節された。 特に”光合成”、”リノール酸m.”、”イソキノリンアルカロイドb.”、”フラボノイドb.”、”デンプンおよびショ糖m”が強く調節されていた(図。 3B)。 最も高い数のDegは、ステージ1のほとんどのペアワイズ比較(1対2は例外であった)、およびステージ2対5の比較(すべての>5,000Deg;表2)で同定された。 はるかに最小の数は、S3とS4の比較(39)で識別されました。 S1対S2のペアでの比較では、’アミノ糖およびヌクレオチド糖m.’が最も高度に特異的に調節された経路であり、続いて’リノール酸m.’が続いた(図1)。 6). S2対S3ペアワイズ比較では、最も有意に差動調節された経路は、’イソキノリンアルカロイドb.’、’チロシンm.’、’フェニルプロパノイドb’、および’フラボノイドb.’であった。 両方のペアワイズ比較では、Degの最大数(それぞれ>15と>35)は、”澱粉とショ糖m”で同定されました。 のS3対… S4の比較では、’(α-)リノール酸m.’が最も有意に調節された経路であったが、遺伝子の数ははるかに低かった。 S4対S5の比較では、’フェニルプロパノイドb’(また、Degの最大数)、および’リノール酸m’が最も有意に調節された経路であった。
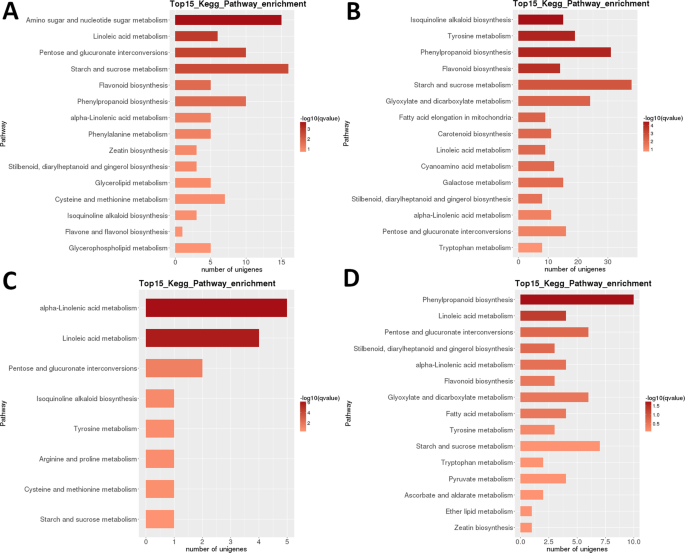
LRトランスクリプトーム:ペアワイズ段階間KEGG代謝経路濃縮。 (A)ステージ1対ステージ2。 (B)ステージ2対ステージ3。 (C)ステージ3対ステージ4。 (D)ステージ4対ステージ5。 上位15(または合計<15の場合はすべて)有意に濃縮された経路が示されている。 バーの色は、q値の大きさを表します(FDR調整されたp値、図に示すカラーチャート)。
LRメタボローム
LRの発達段階のペアワイズ比較における差動調節代謝産物の数は、66(ステージ3対4)から133(ステージ2対5)まで変化した(補足データセットS3)。 連続した段階の比較では、第1段階と第2段階の間で最大の数が観察された(117)。 これらは多数の経路に割り当てられました; “二次代謝産物のb”に割り当てられた代謝産物の数が最も多く、続いて”タンパク質消化吸収”、”アミノ酸のb”、”フラボノイドb”が続く。 S2対S3の比較では、”ABCトランスポーター”が最も有意に濃縮された経路であり、続いて”プリンm”が続いた。 S3対S4ペアの比較では、”多様な環境における微生物m.”が最も有意に濃縮された経路であり、続いて”カルバペネムm.”が続いた。 のS4対… S5対の比較では、比較的多数の経路が同様の結果(2つの代謝産物および類似のp値)を示したが、注目すべきは”イソフラボノイドb”および”フラボノイドb”の出現である(図。 7;補足図)。

LR metabolomicデータ:pairwise interstage KEGGの新陳代謝の細道の強化。 (A)ステージ1対ステージ2。 (B)ステージ2対ステージ3。 (C)ステージ3対ステージ4。 (D)ステージ4対ステージ5。 リッチ因子は、経路における有意に調節された代謝産物の数と、その経路において注釈された代謝産物の総数との比である(範囲=0から1.0)。 点のサイズは、対応する経路における有意に濃縮された代謝産物の数を表し、点の色はP値を表す(図に示す両方の凡例)。
個々の代謝産物(補足データセットS3)に関しては、s1対 S2の比較では、S1でアップレギュレートされた代謝産物のリストは、トレハロース、ガラクチノールおよびL-リンゴ酸(≧19-21log2fc)によって突破されたが、オレイン酸、2-オキソアジピン酸およびステアリン酸は、S2(≧15-18log2fc)で最も高度にアップレギュレートされた代謝産物であった。 S2対S3の比較では、ジヒドロキシアセトン、インドキシル硫酸塩とN-アセチルラクトサミンは、S2(≧17-19.5log2fc)、およびトレハロース、ガラクチノールとL-リンゴ酸(≧19-21log2fc)S3で最も高度にアップレギュレートされた代謝産物であった。 のS3対… S4比較、L-リンゴ酸、DL-アルギニンとオレイン酸は、S3(≧16-19log2fc)、および1,7-ジメチルキサンチン、D-マンノースとN-アセチルラクトサミン(≧15-17log2fc)S4でアップレギュレートされた。 S4対S5の比較では、フラビンモノヌクレオチド、トレハロースとイソフェル酸は、PG(16:0/18:1(9Z))、D-プロリン、およびDL-アルギニンは、最も高度にアップレギュレート(すべての≥16から18log2fc)代謝産物であったのに対し、S4(≥18-20log2fc)でアップレギュレートされた。
異なる発達段階におけるDegの種間比較分析
果実の発達中の総Deg
種間ペアワイズ段階比較(LR1対LB1、LR2対LB2など。 は、9 2 8度が全ての5つの対によって共有されたことを示す(図1 0B)。 8A)。 Degの最大数は、第3段階(3 9 8 9)で同定され、第4段階(2 8 2 5)で最も低かった(図3)。 対に特有の最大数のDegは、第3段階(5 7 4)、第1および第5段階(共に5 5 4)で観察され、第4段階(1 2 6)で最低であった(図8B);対に特有の最大数のDegは、第3段階(5 7 4)、第1およ 8A)。 上方調節度および下方調節度の数は、ペアワイズ段階の比較のそれぞれにおいて比較的類似していた;例えば、第5段階では、LBと比較して、LRでは、1 6 6 8度が、上方調節され、および、1 6 7 0度が下方調節された(図1 0A)。 8B)。 しかし、他の四つの段階では、アップレギュレートされた遺伝子の数はわずかに(93-189度)高かった。

L.barbarum(L b)とl.ruthenicum(L r)の果実間で差動的に発現した遺伝子(Degs)。 (A)詳細な段階的(1〜5)比較(LB対LR)。 (B)五つの研究された発達段階におけるLBと比較して、LRのアップレギュレート(赤)およびダウンレギュレート(緑)度の数。
Transcriptome-pathways
Degのヒートマップ解析は、二つの種の果実は、すべての発生段階で非常に異なる遺伝子発現プロファイルを示すことを示していますが、生物学的複製は、各発生段階で個々の変動の限られた量を示す、非常に類似したプロファイルを示しました(補足図:図。 S9)。 KEGG経路濃縮の比較分析は、全ての5つの発生段階を通して、LBと比較して、LRにおいて、いくつかの経路のみが一貫して高度に濃縮された(遺伝子調節の点 9). 特に、植物ホルモンシグナル伝達(2nd-S1で最高、8th-S2で最高、S3、S4およびS5で最高)と植物-病原体相互作用(S1で最高、3rd-S2で最高、2nd-S3で最高、4th-S4で最高、および15th-S5で最高)は、すべての段階で比較的高度にアップレギュレートされていた。 フェニルプロパノイド生合成(ないトップ15S1で、S2で最高、3位-S3で最高、7位-S4で最高、11位-S5で最高)、ユビキノンおよび他のテルペノイド-キノン生合成(ないトップ15S1で、6位-S2で最高、7位-S3で最高、2位-S4で最高、6位-S5で最高)も、最初のものを除くすべての段階で比較的高度にアップレギュレートされた。 フラボノイド生合成経路は、初期段階で高度に濃縮されていなかった(s1のトップ15ではなく、S2のトップ14-最高)、および後期段階で高度に濃縮されていた(3-4-ステージ3-5の間に最高)。 (α-)リノール酸代謝は、中間段階で高度に濃縮されていた(9th-S1で最高、2nd-S2で最高、5thと6th-S3で最高、8th-S4で最高、s5でトップ15ではない)。

KEGG代謝経路濃縮の比較分析。 L.barbarumと比較してL.ruthenicumに富むトップ15の経路が左(赤)に示され、l.barbarumと比較してl.ruthenicumに富む経路が右(緑)に示されている。 発達段階(1-5)が図に示されている。 q値は、FDR調整されたp値です。
l.barbarumと比較して、L.ruthenicumでダウンレギュレートされた経路の中で(図10)。 9)注目すべき変化は、初期段階(1と2)、シアノアミノ酸代謝とカロテノイド生合成が最も高度にダウンレギュレートされたとき、後期段階(4と5)、小胞輸送、ニコチン酸とニコチンアミド代謝、ポルフィリンとクロロフィル代謝におけるSNARE相互作用が一貫して比較的高度にダウンレギュレートされたときに観察された。
トランスクリプトーム–個々の遺伝子
最も高度に差動発現された遺伝子のうち、いくつかは発達段階特異的であった(すなわち、発達段階の初期または後期にのみ高度に差動的に調節された)が、いくつかは研究された五つの段階全体にわたって一貫して高度に差動的に調節されていた(補足データセットS4)。 いくつかの免疫関連遺伝子は非常に非常に初期の発達段階でLBと比較してLRでアップレギュレートされ、発達段階特異的発現パターンの例の一つです: キチナーゼは、S1(13.43倍)で2番目に高いupregulated DEG、S2(13.89倍)で最も高かったが、後の段階ではDEGではなかった。 同様に、EIX受容体1/2も初期段階で非常に高度にアップレギュレートされ、S1(13.70)で最も高く、S2(10.90)で2番目に高いが、後の段階ではDEGとしても同定されなかった。 いくつかの成長関連遺伝子も同様の発現パターンを示した:ホスホグリセリン酸キナーゼ(PGK)は、最初の三つの段階(13.14、12.87と12)で最も高度にアップレギュレー77)であったが、後の段階ではDEGと同定されていなかった。 CCR4-ない転写複合体サブユニット7/8(CNOT7/8)も非常に同様の発現パターンを示した:高度に最初の三つの段階でアップレギュレートし、ステージ4と5のDEGではな いくつかのフラボノイドおよびフェニルプロパノイド生合成関連遺伝子は、初期段階では比較的低い発現を有し、後の段階では非常に高い、逆発達期特 例としては、二官能性ジヒドロフラボノール4-レダクターゼ/フラバノン4-レダクターゼ(DFR)があり、これはS1(2)でLRでわずかにアップレギュレートされていた。25)、S2のDEGではなく、s3(7.79)で高度にアップレギュレートされ、S4(14.25)およびS5(16.03)で3番目に高いアップレギュレートされた遺伝子。 この遺伝子のパラローグは、ほぼ同じパターンを示した:わずかにS1(2.44)ではなく、S2のDEG、高度にS3(7.40)でアップレギュレート、6番目に高いアップレギュレートDEG S4(13.26)と5番目のS5(14.59)でアップレギュレート。 同様に、フラボノイド3’、5′-ヒドロキシラーゼ(F3’5’H)は、高度にS3(6.69)、S4(13.42)で5番目に高いupregulated遺伝子、およびS5(15.05)で4番目に高いUPREGULATED最初の二段階でDEG フラボノイドO-メチルトランスフェラーゼ(OMT)は、S1ではDEGではなかったが、S2ではすでに中程度の高アップレギュレーションレベル(4.32)を示し、S3ではすでに三番目に高いアップレギュレーションされたDEG(13.30)であり、S4(18.73)とS5(18.10)で最も高いアップレギュレーションされた遺伝子であった。 ロイコアントシアニジンジオキシゲナーゼ(LDOX;アントシアニン生合成)は、s1とS2のDEGではなく、後の段階(それぞれ5.63、9.44、11.56)で非常に高いアップレギュレーショ 二つのカルコンシンターゼパラログ(CHSとCHS2;フラボノイド生合成)も高度にS1とS2(CHS2:ないDEG、CHS)で調節されていませんでした: -1.14S1ではなく、S2でDEG)が、S3–S5では両方の遺伝子が中-高から高までのアップレギュレーションを示した(CHS2:5.32、7.84、6.00;およびCHS:4.67、7.01、6.82;それぞれ)。 これらの遺伝子をqPCR解析のために選択し、結果はRNA-seqデータ(補足結果;補足データセットS5)と非常に一致しています。 最後に、サイトカイニンデヒドロゲナーゼ、ゼアチン生合成関連遺伝子は、また、ますます最後の三つの段階(2.6-5.7)の間にアップレギュレートされました。
しかし、いくつかの遺伝子は、研究された五つの段階すべてを通じて一貫して差動的に発現していた。 9.38と8.58(それぞれの順序で倍変化として提示されるすべての値)S1、6.30と6.34S2、2番目と7番目に高いアップレギュレートDeg S3(14.08と12.70)、2番目と4番目に高いS4(15.71と6.34)である。14.16)とs5で2番目と6番目の最高(16.48と14.40)。 植物病害抵抗性タンパク質RPM1も、全ての5段階で高度にアップレギュレートされた(S1=1 3:1 5;S2=1 2:0 8;S3=1 3:1 1、S4=1 2:8 1;S5=1 3:9 4)。 すべての発達段階を通じて一貫して差動発現遺伝子の中でも、いくつかは、アミノ酸代謝に関連していたが、そのパターンは、以前の例と比較して逆転した:彼らはLBと比較してLRで高いダウンレギュレーションを示した。 例はアセチルCoAアシルトランスフェラーゼ1(AAT1;バリン、ロイシンおよびイソロイシン分解)であり、s1の−7.0から<-10倍の最後の三つの段階で始まる、ますます高いダウンレギュレーションの時間的プロファイルである。 プロリンイミノペプチダーゼは、アルギニンとプロリン代謝に関連付けられている、非常にすべての段階でLRでダウンレギュレートされた:S1=-9.75、S2=-10.89(3rd-高)、S3=-11.05(4th-高)、S4=-10.01、およびS5=-11.98(3rd-高)。 最後に、5–methyltetrahydropteroyltriglutamate-ホモシステインメチルトランスフェラーゼ(metE)は一貫して非常に高度にすべての段階でLRでダウンレギュレートされた:2番目に高いS1(-11.76)、S2(-11.74)、3番目に高いS3(-11.43)、S4(-12.36)、および2番目に高いS5(-12.83)。 GTP結合核タンパク質Ran(RAN;-10.0–12.0)と複製因子A1(RFA1)(-8.0–12.0):二つのDNA複製と転写関連遺伝子も非常にすべての段階でLRでダウンレギュレートされた。 いくつかの成長とストレス関連遺伝子も一貫して非常にLRでダウンレギュレートされた:異種核リボ核タンパク質A1/A3(hnRNP;-7-11)と熱ショック70kDaタン 興味深いことに、フェニルプロパノイド生合成関連遺伝子、シキメートヒドロキシシンナモイルトランスフェラーゼ(HCT)は、また、一貫して非常にLRでダウンレギュレートされた:S1=-6.82、S2=-8.14、S3=-11.71(最高)、S4=-11.00(3番目に高い)、S5=-11.91(4番目に高い)。 しかし、アントシアニン生合成、転写因子MYB114のキーレギュレータは、非常にすべての五つの発達段階の間にLRでアップレギュレートされた:6.11、4.69、7.47、9.05、および8.95(S1–S5
メタボローム–経路
我々はまた、代謝経路の比較種間段階的解析を行った(図。 10). 最初の発達段階(S1)では、我々は39差動調節代謝産物を同定した。 これらの代謝産物が関連していた上位20の経路のうち、それらのいくつかはアミノ酸に関連していたが、経路ごとの代謝産物の総数は比較的小さく(1-2)、P値は高いレベルの有意性を示唆していなかった(図。 10-パネル1)。 特にビタミンB6代謝(m)と多様な環境での微生物mは、両方の比較的高いP値、1.0の濃縮因子(EF)、および2同定された代謝産物を示した。 S2では、トリプトファンm、フェニルプロパノイド生合成(b)、フェニルプロパノイドのb(これらはKEGGデータベース内の二つの異なる経路である)、およびフェニルアラニン、チロシンおよびトリプトファンb(すべてのEF=1.0、2-3代謝産物、およびp<0.5;図。 10-パネル2)。 S3において、本発明者らは、19の経路に関連する59の差動調節代謝産物を同定し、ほとんどすべてがEF1.0であるが、比較的有意ではないP値(>0.5;図 10-パネル3)。 代謝産物の比較的高い数(n=5)を有する経路は、タンパク質の消化および吸収、植物の二次代謝産物のb、抗生物質のb、およびアミノ酸のbであった。 S4において、本発明者らは、多数の経路、主にEF1.0、および比較的高い有意性値(主にP>0.5;図5)に関連する58の特異的に調節された代謝産物を同定した。 10-パネル4)。 代謝産物の数が比較的多い経路(n≥3)は、フェニルプロパノイドb、フェニルアラニン、チロシンおよびトリプトファンb、グルコシネートb、シキメート経路に由来するアルカロイドのb、および2-オキソカルボン酸mであった。熟した果実(S5)では、39の異なる調節された代謝産物を同定し、多数の経路に関連しているが、主にp値が低く、経路ごとに1つの代謝産物のみである。 10-パネル5)。 1つ以上の代謝産物を持つ経路は、次のとおりであった。: すべてのデータ(2種×5段階×5生物学的複製)の主成分分析(PCA)は、生物学的複製(クラスタリング)の間で高い類似性を明らかにし、両方の種の異なる果実熟成期間の顕著な変動を確証した(図)。 10-パネル6)。

KEGG代謝経路濃縮の比較分析。 L.barbarumと比較してL.ruthenicumに富むトップ15の経路が左(赤)に示され、l.barbarumと比較してl.ruthenicumに富む経路が右(緑)に示されている。 発達段階(1-5)が図に示されている。 q値は、FDR調整されたp値です。
メタボローム-個々の代謝産物
二つの種の間で最も高度に差動調節された代謝産物のリスト(補足データセットS6)は、五つの果実の発達段階の間でいく 興味深いことに、フルクトース1-リン酸は、すべての五つの段階の間に、LBと比較して、LRの最も高度にupregulated代謝産物であった:log2倍の変化=6.3、7.6、7.7、8.1、および6.5( LBで非常にupregulated代謝物質に関しては、段階間のより多くの変化がありました: S1では、違いは最も高度にupregulated代謝産物として9-デセノールで、かなり小さかった(log2倍の変化=2.7;LRと比較して)。 S2とS3の分析は、最も高度にupregulated代謝産物としてフェノール(それぞれ3.7と3.2)と、非常に合同な結果を生成しました。 S4では、インドキシル硫酸塩(4.7)はLBでupregulatedトップ代謝産物であった。 代謝シフトは、LBでupregulated代謝産物のリストは、ステアロイルカルニチン(7.1)、メトキシ酢酸(5.3)、S-メチル-5′-チオアデノシン(4.7)、リシノプリル(4によって突破された熟した果実(S5)で観察された。7)、アデノシン3’、5′-環状リン酸(cAMP)(4.7)など。 LRで高度にアップレギュレートされた他の代謝産物(フルクトース1-リン酸から離れて)は、ナリンギン(6.2)、ラウロイルCoA(4.8)、L-プネイルアラニン(4.6)などであった。
« バルディビア