vi samlet frukt Av L. barbarum og L. ruthenicum på fem utviklingsstadier, fra ung frukt (≈10 dager etter blomstring) til moden (moden) frukt (34-45 dager etter blomstring), og studerte deres transkriptome og metabolom.
- rna-seq de novo montering og funksjonell merknad av unigenes
- Lycium barbarum (LB): parvise interstage analyser
- Lb Transkriptome
- Lb Metabolom
- Lycium ruthenicum (LR): parvise interstage analyser
- Lr Transkriptom
- Lr-Metabolom
- Interspesifikk komparativ analyse Av DEGs på forskjellige utviklingsstadier
- Total DEGs under fruktutviklingen
- Transcriptome-pathways
- Transkriptom – individuelle gener
- Metabolome-pathways
- Metabolom-individuelle metabolitter
rna-seq de novo montering og funksjonell merknad av unigenes
vi forberedte totalt 30 cDNA-biblioteker fra frukt Av L. barbarum og L. ruthenicum, med tre biologiske replikater (tre frukter fra tre trær) ved hvert tidspunkt: 2 arter × 5 tidspunkter × 3 biologiske replikater. Prøver ble merket LB/LR(1-5)-(1-3), HVOR LB Er L. barbarum og LR Er l. ruthenicum, 1-5 er utviklingsstadier av frukt (S1–S5), og 1-3 er individuelle prøver (biologiske replikater); så FOR eksempel LB1-1 representerer L. barbarum, 1. samplet utviklingsstadium (S1), fruktprøve nr. 1 (av tre). Vi genererte over 1.72 milliarder par-end leser for disse 30 cDNA-bibliotekene, tilsvarende et gjennomsnitt på 57.2 millioner leser per prøve (Supplerende Datasett S1). Streng kvalitetsvurdering og datafiltrering ga totalt 801.766 høykvalitets leser med gjennomsnittlig lengde på 730 og N50 på 1107 bp (Tabell 1). Til slutt ble totalt 326 276 unigener med gjennomsnittlig lengde på 596 bp og N50 av 847 bp hentet fra transkripsjonene (Tabell 1). Korrelasjonskoeffisienter FOR rna-seq-data for de 30 prøvene indikerer meget god konsistens av resultater blant biologiske replikater (Fig. 2).

Heatmap av korrelasjonskoeffisienter FOR RNA-seq data for 30 prøver Av L. barbarum (LB) og L. ruthenicum (LR) frukt på fem forskjellige utviklingsstadier. Prøver er merket LB / R1–5_1-3, HVOR LB er l. barbarum, LR er L. ruthenicum, 1-5 er frukt utviklingsstadier, og 1-3 individuelle prøver. Prøver ble gruppert etter hierarkisk clustering; dendrograms over og til venstre for varmekartet indikerer slektskap av prøver.
blant alle 326.276 unigener som ble spurt mot offentlige databaser, ble totalt 193.021 (59.15%) matchet gener og/eller proteiner i minst en database, og 12.171 (3.73%) annotert i alle databaser. Det største antallet unigener (149 863, 45,93%) ble merket i nt-databasen, og det laveste antallet (24 017; 7,36%) i kog-databasen.
Lycium barbarum (LB): parvise interstage analyser
Lb Transkriptome
det høyeste antallet DEGs ble identifisert i alle parvise sammenligninger av 1. trinn, og i 2. vs 5. trinns sammenligning (alle > 10 000 degs; Tabell 2). De minste tallene ble identifisert i 3. vs 4. og 4. vs 5. trinn sammenligninger (255-257). Heatmap analyse Av DEGs i LB viser at ganske forskjellige sett av gener var svært oppregulert i de tidlige utviklingsstadiene (1 + 2) og i senere stadier (3 til 5) (Fig. 3A). Sample relatedness analyse indikerer at prøvene kan deles inn i to klader (trinn 1 + 2 og 3 + 4 + 5), med sistnevnte clade videre delt inn i to clades: trinn 3 + 4 og trinn 5. Intraspesifikk kegg funksjonell klassifiseringsanalyse av Disse DEGs identifiserte 15 veier signifikant (P < 0,05) differensielt regulert mellom ulike utviklingsstadier (Supplerende Datasett S2). Spesielt sterkt differensielt regulert var ‘plantehormon signaltransduksjon’, ‘ fenylpropanoid biosyntese (b.)’, ‘ linolsyre metabolisme (m.)’, ‘stivelse og sukrose m.’, og ‘ zeatin b. ‘(Fig. 3A).

Heatmaps og funksjonelle pathway analyser av differensielt uttrykte gener (DEGs) I Lycium barbarum (panel A) og L. ruthenicum (panel B) frukt. Heatmaps ble generert av en hierarkisk analyse Av DEGs (y-akse) og individuelle prøver (x-akse), hvor dendrograms over og til venstre for heatmap indikerer slektskap av prøver. Prøvene er merket LB / R_1–5_1-3, hvor arten akronym (LB eller LR) etterfølges av utviklingsstadiet av frukt (1-5), og prøvenummeret (1-3). Intraspesifikke kegg pathway analyser Av DEGs i alle fem utviklingsstadier i de to artene er vist til høyre for heatmaps. Bare de 15 beste anrikede banene er oppført. q-verdi er EN fdr-justert p-verdi.
For en mer grundig analyse av dataene fokuserte vi på sammenligning av mest signifikant regulerte veier i suksessive utviklingsstadier. I den første parvise sammenligningen (S1 vs S2) var ‘fenylpropanoid b.’ den mest differensielt regulerte banen, etterfulgt av ‘stivelse og sukrose m.’ (Fig. 4). Svært stort Antall DEGs (> 100) ble identifisert i begge veier. Et lignende resultat ble observert i følgende parvis sammenligning, S2 vs S3, men til tross for ganske stort antall degs (>80) ‘stivelse og sukrose m. viste noe lavere q-verdi. I S3 vs. S4 sammenligning,’ karbonfiksering i fotosyntetiske, organismer ‘ var den mest betydelig regulert vei, men antall gener var mye lavere. I det siste paret var S4 vs S5, ‘zeatin b.’, flavonoid b.’, fettsyre b.’ og ‘galaktose m.’ de mest regulerte banene, men ingen av banene viste mer enn to Grader.

lb transcriptome: parvis interstage kegg metabolisk vei berikelse. Trinn 1 vs. trinn 2. (B) Trinn 2 vs. trinn 3. (C) Trinn 3 vs. trinn 4. (D) Trinn 4 vs. trinn 5. Topp 15 (eller alle hvis totalt <15) betydelig beriket baner vises. Fargen på linjen representerer størrelsen på q-verdien (EN fdr-justert p-verdi). Fargekart er vist i figuren.
Lb Metabolom
vi gjennomførte parvise sammenligninger av utviklingsstadier for å identifisere anrikning av metabolitter mellom alle trinnspar. Antallet differensielt regulerte metabolitter i parvise sammenligninger av utviklingsstadier i LB varierte fra 66 (trinn 3 vs 4) til 129 (trinn 2 vs 5). Det største tallet i suksessive trinnsammenligninger ble observert mellom 1.og 2. trinn (117). Disse ble tildelt et stort antall veier; med det største antallet metabolitter tildelt ‘ b. av sekundære metabolitter ‘(også den laveste p-verdien), etterfulgt av ‘protein fordøyelse og absorpsjon’ og ‘ b. av aminosyrer ‘(Fig. 5; Utfyllende Tall). I den påfølgende fasen par sammenligning (S2 vs. S3),’ ABC transportører ‘var den mest betydelig beriket vei, etterfulgt av’ purine m.’. I s3 vs S4 par sammenligning, ‘mikrobiell m. i ulike miljøer’ var den mest betydelig beriket vei, etterfulgt av ‘ carbapenem m.’. I s4 vs S5 par sammenligning, et relativt stort antall veier viste lignende resultater (to metabolitter og lignende p-verdier), men bemerkelsesverdig er utseendet på ‘isoflavonoid b.’ og ‘flavonoid b.’ blant dem (Fig. 5; Utfyllende Tall).

lb metabolomiske data: parvis interstage kegg metabolisk vei berikelse. Trinn 1 vs. trinn 2. (B) Trinn 2 vs. trinn 3. (C) Trinn 3 vs. trinn 4. (D) Trinn 4 vs. trinn 5. Rich factor er forholdet mellom antall signifikant regulerte metabolitter i banen og totalt antall metabolitter merket i den banen (område = 0 til 1,0). Størrelsen på punktet representerer antall signifikant berikede metabolitter i den tilsvarende banen, og fargen på punktet representerer P-verdien (begge legender vist i figuren).
når det gjelder individuelle metabolitter, i S1 vs S2-sammenligningen, ble listen over metabolitter oppregulert I S1 toppet Av Trehalose, Glaktinol og L-Eplesyre(alle ≈20 log2FC). Oljesyre, 2-Oksoadipinsyre og Stearinsyre var de høyest oppregulerte metabolittene i S2 (alle ≈15 til 18 log2FC). Listen over høyest oppregulerte metabolitter i s2 sammenlignet Med S3 ble toppet Med Dihydroksyaceton, LysoPC(18:1(9Z)) og Adenin (alle ≈16 til 17,5 log2FC). Listen over de mest oppregulerte metabolittene I S3 (sammenlignet Med S2) ble toppet Av Trehalose, Galaktinol og L-Eplesyre(alle ≈19 til 20 log2FC). I s3 vs. S4-sammenligningen ble listen over de mest oppregulerte metabolittene I S3 toppet Med L-Eplesyre, DL-Arginin og Oljesyre (alle ≈18 til 19 log2FC), mens listen over de mest oppregulerte metabolittene I S4 ble toppet Med D-Mannose, N-Acetyllaktosamin og LysoPC(18:1(9Z)) I LB (alle ≈16 log2FC). I S4 vs. Til sammenligning ble listen over de mest oppregulerte metabolittene i s4 toppet Av Trehalose, Palmitinsyre, N-Acetyllaktosamin (alle ≈17 til 18 log2FC) i LB. Listen over de mest oppregulerte metabolittene I S5 ble toppet Med L-Norleucin, Antranilsyre (Vitamin L1) og DL-Arginin (alle ≈16 til 18 log2FC) (Supplerende Datasett S3).
Lycium ruthenicum (LR): parvise interstage analyser
Lr Transkriptom
Heatmap analyse Av degs i LR viser at nesten helt forskjellige sett av gener var svært oppregulert i de tidlige utviklingsstadiene (1 + 2) og i moden frukt (trinn 5), med en tilsynelatende transkriptomisk tilbakestilling som oppstår etter andre trinn (Fig. 3B). Sample relatedness analyse indikerer at prøvene kan deles inn i to klader (trinn 1 + 2 og 3 + 4 + 5), med sistnevnte clade videre delt inn i to clades: trinn 3 + 4 og trinn 5. Intraspesifikk kegg funksjonell klassifiseringsanalyse av Disse DEGs identifiserte 35 veier signifikant (P < 0,05) differensielt regulert mellom ulike utviklingsstadier (Supplerende Datasett S2). Spesielt sterkt differensielt regulert var ‘fotosyntese’, ‘ linolsyre m.’, ‘ isokinolinalkaloid b.’, ‘ flavonoid b. ‘og’ stivelse og sukrose m ‘ (Fig. 3B). Det høyeste antallet DEGs ble identifisert i de fleste parvise sammenligninger av trinn 1 (1 vs. 2 var et unntak), og i trinn 2 vs. 5 sammenligning (alle >5000 degs; Tabell 2). Langt det minste tallet ble identifisert I s3 vs S4-sammenligningen (39). I S1 vs S2 parvis sammenligning, ‘amino sukker og nukleotid sukker m.’ var den mest differensielt regulert vei, etterfulgt av ‘linolsyre m.’ (Fig. 6). I s2 vs S3 parvis sammenligning var de mest signifikant differensielt regulerte banene ‘ isokinolinalkaloid b.’, ‘ tyrosin m.’, ‘fenylpropanoid b’ og ‘ flavonoid b.’. I begge parvise sammenligninger ble det høyeste antallet DEGs (henholdsvis>15 og >35) identifisert i ‘stivelse og sukrose m.’. I S3 vs. S4 sammenligning,’ (alfa-)linolsyre m. ‘ var den mest signifikant regulerte banen, men antall gener var mye lavere. I s4 vs S5-sammenligningen var’ fenylpropanoid b ‘(også det største antallet DEGs) og’ linolsyre m. ‘ de mest regulerte veiene.
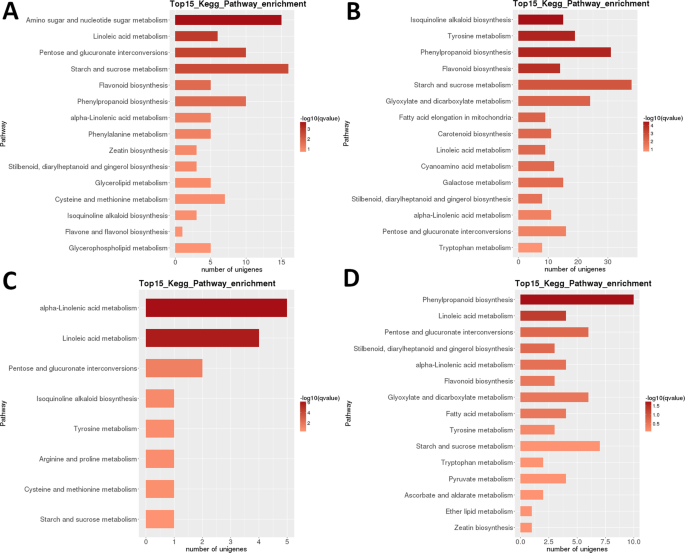
lr transcriptome: parvis interstage kegg metabolisk vei berikelse. Trinn 1 vs. trinn 2. (B) Trinn 2 vs. trinn 3. (C) Trinn 3 vs. trinn 4. (D) Trinn 4 vs. trinn 5. Topp 15 (eller alle hvis totalt <15) betydelig beriket baner vises. Fargen på linjen representerer størrelsen på q-verdien (EN FDR-justert p-verdi, fargediagram vist i figuren).
Lr-Metabolom
antall differensielt regulerte metabolitter i parvise sammenligninger av utviklingsstadier i LR varierte fra 66 (trinn 3 vs 4) til 133 (trinn 2 vs 5) (Supplerende Datasett S3). I suksessive trinnsammenligninger ble det største tallet observert mellom 1.og 2. trinn (117). Disse ble tildelt et stort antall veier; med det største antallet metabolitter tildelt ‘b av sekundære metabolitter’, etterfulgt av ‘protein fordøyelse og absorpsjon’, ‘b. av aminosyrer’, og’flavonoid b’. I s2 vs. S3 sammenligningen, ‘ABC transportører’ var den mest betydelig beriket vei, etterfulgt av ‘purine m’. I s3 vs S4 par sammenligning, ‘mikrobiell m. i ulike miljøer’ var den mest betydelig beriket vei, etterfulgt av ‘ carbapenem m.’. I S4 vs. S5 par sammenligning, et relativt stort antall veier viste lignende resultater (2 metabolitter og lignende p-verdier), men bemerkelsesverdig er utseendet av ‘isoflavonoid b.’ og ‘flavonoid b.’ blant dem (Fig. 7; Utfyllende Tall).

LR metabolomic data: parvis interstage kegg metabolsk vei berikelse. Trinn 1 vs. trinn 2. (B) Trinn 2 vs. trinn 3. (C) Trinn 3 vs. trinn 4. (D) Trinn 4 vs. trinn 5. Rich factor er forholdet mellom antall signifikant regulerte metabolitter i banen og totalt antall metabolitter merket i den banen (område = 0 til 1,0). Størrelsen på punktet representerer antall signifikant berikede metabolitter i den tilsvarende banen, og fargen på punktet representerer P-verdien (begge legender vist i figuren).
når det gjelder individuelle metabolitter (Supplerende datasett S3), i S1 vs. S2 sammenligning ble listen over metabolitter oppregulert i S1 toppet Av Trehalose, Galaktinol og L-Eplesyre (≈19-21 log2FC), Mens Oljesyre, 2-Oksoadipinsyre og Stearinsyre var de mest oppregulerte metabolittene I S2 (≈15-18 log2FC). I s2 vs. S3-sammenligningen var Dihydroksyaceton, Indoksylsulfat og N-Acetyllaktosamin mest oppregulerte metabolitter i s2 (≈17-19, 5 log2FC) og Trehalose, Galaktinol Og L-Eplesyre (≈19-21 log2FC) I S3. I S3 vs. S4 sammenligning Ble L-Eplesyre, DL-Arginin og Oljesyre oppregulert i S3 (≈16-19 log2FC), og 1,7-Dimetylxantin, D-Mannose og N-Acetyllaktosamin (≈15-17 log2FC) I S4. I s4 vs. S5-sammenligningen ble Flavinmononukleotid, Trehalose og Isoferulsyre oppregulert i s4 (≈18-20 log2FC), MENS pg(16:0/18:1(9Z)), D-Prolin og DL-Arginin var mest oppregulert (alle ≈16 til 18 log2FC) metabolitter.
Interspesifikk komparativ analyse Av DEGs på forskjellige utviklingsstadier
Total DEGs under fruktutviklingen
Interspesifikk parvis trinns sammenligning (LR1 vs LB1, LR2 vs LB2, etc.) viser at 928 DEGs ble delt av alle fem parene (Fig. 8A). Det høyeste antallet DEGs ble identifisert i trinn 3 (3989), og det laveste i trinn 4 (2825) (Fig. 8B); mens det høyeste antallet DEGs unike for et par ble observert i trinn 3 (574), 1 og 5 (begge 554), og det laveste i trinn 4 (126) (Fig. 8A). I trinn 5 ble 1668 DEGs oppregulert og 1670 DEGs ble nedregulert I LR i forhold TIL LB (Fig . 8B). Men i de andre fire stadiene var antall oppregulerte gener litt (93 til 189 Grader) høyere.

Gener differensielt uttrykt (DEGs) mellom frukt Av L. barbarum (LB) og L. ruthenicum (LR). (A) Detaljert stagewise (1 til 5) sammenligninger(LB vs LR). (B) antall oppregulerte (røde) og nedregulerte (grønne) Grader I LR i forhold TIL LB i fem studerte utviklingsstadier.
Transcriptome-pathways
Heatmap analyse Av DEGs indikerer at fruktene av to arter viser svært forskjellige genuttrykk profiler under alle utviklingsstadier, men biologiske replikater utstilt svært like profiler, noe som indikerer en begrenset mengde individuell variasjon i hvert utviklingsstadium (Supplerende Tall: Fig. S9). Komparativ analyse av kegg pathway berikelse viser at bare noen veier var konsekvent høyt beriket (i form av genregulering)I LR i forhold TIL LB gjennom alle fem utviklingsstadier (Fig. 9). Spesielt, plante hormon signaltransduksjon (2nd-høyeste I S1, 8th-høyeste I S2, den høyeste I S3, S4 Og S5) og plante-patogen interaksjon (den høyeste I S1, 3rd-høyeste I S2, 2nd-høyeste I S3, 4th-høyeste I S4, og 15th-høyeste I S5) var relativt høyt oppregulert i alle stadier. Fenylpropanoidbiosyntese (ikke i topp 15 I S1, den høyeste I S2, 3.høyeste I S3, 7. høyeste I S4, 11. høyeste I S5), ubiquinon og annen terpenoid-kinonbiosyntese (ikke i topp 15 I S1, 6. høyeste I S2, 7. høyeste I S3, 2. høyeste I S4, 6. høyeste I S5) var også relativt høyt oppregulert i alle stadier unntatt den første. Flavonoid biosyntese pathway var ikke sterkt beriket i tidlige stadier (ikke i topp 15 I S1, 14th-høyest I S2), og svært beriket i sent stadier (3.til 4.-høyest i trinn 3 til 5). (alfa-)Linolsyre metabolisme var svært beriket i mellomstadier (9. – høyest I S1, 2. – høyest I S2, 5. og 6.-høyest I S3, 8. – høyest I S4, ikke i topp 15 I S5).

Komparativ analyse AV kegg metabolisk vei berikelse. De 15 beste banene beriket Med L. ruthenicum i forhold Til l. barbarum er vist til venstre (rød), og de som er beriket Med L. barbarum i forhold Til l. ruthenicum til høyre (grønn). Utviklingsstadier (1-5) er angitt i figuren. q-verdi er EN fdr-justert p-verdi.
blant veiene nedregulert I L. ruthenicum i forhold Til L. barbarum (Fig. 9) merkbare endringer ble observert mellom de tidlige stadiene (1 og 2), da cyanoaminsyremetabolismen og karotenoidbiosyntesen var mest nedregulert, og sene stadier (4 og 5), da SNARE-interaksjoner i vesikulær transport, nikotinat-og nikotinamidmetabolismen og porfyrin-og klorofyllmetabolismen var konsekvent relativt høyt nedregulert.
Transkriptom – individuelle gener
blant de mest differensielt uttrykte gener var noen utviklingsstadiumspesifikke (dvs. svært differensielt regulert bare i tidlige eller sene utviklingsstadier), men noen var konsekvent svært differensielt regulert gjennom alle fem studerte stadier (Supplerende Datasett S4). Flere immunitetsrelaterte gener svært høyt oppregulert I LR i forhold TIL LB i de tidlige utviklingsstadiene er blant eksemplene på utviklingsstadiumspesifikke uttrykksmønster: chitinase var den 2. høyeste oppregulerte DEG i S1 (13,43 ganger), den høyeste I S2 (13,89 ganger), men i senere stadier var DET ikke EN DEG. TILSVARENDE VAR EIX-reseptor 1/2 også svært oppregulert i tidlige stadier, den høyeste I S1 (13,70) og 2.-høyeste I S2 (10,90), men DET ble heller ikke identifisert SOM DEG i senere stadier. Noen vekstrelaterte gener viste også et lignende uttrykksmønster: fosfoglyseratkinase (PGK) var blant de håndfull mest oppregulerte gener i de tre første stadiene (13.14, 12.87 og 12.77), men DET ble heller ikke identifisert SOM DEG i senere stadier. CCR4-IKKE transkripsjon kompleks underenhet 7/8 (CNOT7/8) viste også et veldig lignende uttrykksmønster: høyt oppregulert i tre første trinn, og ikke EN DEG i trinn 4 og 5. Flere flavonoid-og fenylpropanoidbiosynteseassosierte gener viste et reversert utviklingsstadiumspesifikt uttrykksmønster, med relativt lavt uttrykk i tidlige stadier og svært høyt i senere stadier. Eksempler er: bifunksjonell dihydroflavonol 4-reduktase / flavanon 4-reduktase (DFR), som ble litt oppregulert I LR I S1 (2 .25), ikke EN DEG I S2, høyt oppregulert I S3 (7.79), og 3. høyeste oppregulerte gen I S4 (14.25) Og S5 (16.03). En paralog av dette genet viste et nesten identisk mønster: litt oppregulert I S1( 2.44), ikke EN DEG I S2, høyt oppregulert I S3 (7.40), 6.høyeste oppregulert DEG I S4 (13.26) og 5. I S5 (14.59). Tilsvarende var flavonoid 3′,5′-hydroksylase (F3’5′ H) ikke EN DEG i de to første stadiene, høyt oppregulert I S3 (6,69), 5.høyeste oppregulerte gen I s4 (13,42) og 4. høyeste I S5 (15,05). Flavonoid o-metyltransferase (LAR) var ikke EN DEG I S1, men I S2 viste den allerede et middels høyt oppreguleringsnivå (4.32), Ved S3 var det allerede det tredje høyeste oppregulerte DEG (13.30), og det var det høyeste oppregulerte genet I S4 (18.73) Og S5 (18.10). Leukoantocyanidindioksygenase (LDOX; anthocyaninbiosyntese) var ikke EN DEG i S1 og S2, etterfulgt av høy til svært høy oppregulering i senere stadier (henholdsvis 5,63, 9,44, 11,56). To chalcone syntase paralogues (CHS OG CHS2; flavonoid biosyntese) ble heller ikke sterkt regulert I S1 Og S2 (CHS2: ikke EN DEG, CHS: -1,14 I S1, ikke EN DEG I S2), men I S3–S5 viste begge gener en middels høy til høy oppregulering (CHS2: 5,32, 7,84, 6,00; og CHS: 4,67, 7,01, 6,82; henholdsvis). Vi valgte disse genene for qpcr-analyse, og resultatene er svært kongruente MED RNA-seq-dataene (Supplerende Resultater; Supplerende Datasett S5). Endelig ble cytokinin dehydrogenase, et zeatin biosynteserelatert gen, også i økende grad oppregulert i løpet av de tre siste stadiene (2,6–5,7).
noen gener ble imidlertid konsekvent differensielt uttrykt gjennom alle fem studerte stadier. Eksempler inkluderte også noen immunitetsrelaterte gener, for eksempel to paraloger av glutation S-transferase, høyt oppregulert I LR i forhold TIL LB i alle stadier: 9.38 og 8.58 (alle verdier presentert som fold-endringer i respektive rekkefølge) I S1, 6.30 og 6.34 I S2, 2.og 7. høyeste oppregulerte DEGs I S3 (14.08 og 12.70), 2. og 4. høyeste I S4 (15.71 og 14,16) og 2. og 6. høyeste i s5 (16,48 og 14,40). Plantesykdomsmotstandsprotein RPM1 ble også sterkt oppregulert i alle fem stadier (S1 = 13,15; S2 = 12,08; S3 = 13,11, S4 = 12,81; S5 = 13: 94). Blant de konsekvent differensielt uttrykte gener gjennom alle utviklingsstadier var også noen relatert til aminosyremetabolismen, men deres mønster ble reversert i forhold til tidligere eksempler: de viste høy nedregulering i LR sammenlignet MED LB. Eksempler er acetyl-CoA acyltransferase 1 (aat1; valin, leucin og isoleucin degradering), med en temporal profil av stadig høy nedregulering, fra -7,0 I S1 til < −10 ganger i de tre siste stadiene. Proliniminopeptidase, assosiert med arginin og prolinmetabolisme, var sterkt nedregulert I LR i alle stadier: S1 = -9,75, S2 = -10,89 (3.høyeste), S3 = -11,05 (4. høyeste), S4 = -10,01 og S5 = -11,98 (3. høyeste). Endelig var 5-metyltetrahydropteroyltriglutamat-homocysteinmetyltransferase (metE) konsekvent ekstremt høyt nedregulert I LR i alle stadier: 2.høyeste I S1 (-11.76), den høyeste I S2 (-11.74), 3. høyeste I S3 (-11.43), den høyeste I S4 (-12.36) og 2. høyeste I S5 (-12.83). To DNA-replikasjon og transkripsjonsassosierte gener ble også sterkt nedregulert I LR i alle stadier: GTP-bindende nukleært protein Ran (RAN; -10,0 til -12,0) og replikasjonsfaktor A1 (RFA1) (-8,0 til -12,0). Noen vekst-og stressrelaterte gener ble også konsekvent sterkt nedregulert i LR: heterogent nukleært ribonukleoprotein A1/A3 (hnrnp; -7 til -11) og varmesjokk 70 kDa protein 1/8 (HSPA1_8) S1 = -4,95, S2 = -8,88, S3 = -11,48 (2.høyeste), S4 = -9,36, s5 = -12,89 (høyest). Interessant nok var et fenylpropanoid biosynteserelatert gen, shikimathydroksykinnamoyltransferase (HCT), også konsekvent sterkt nedregulert I LR: S1 = -6,82, S2 = -8,14, S3 = -11,71 (den høyeste), S4 = -11,00 (3.høyeste), S5 = -11,91 (4. høyeste). En viktig regulator av anthocyaninbiosyntese, transkripsjonsfaktor MYB114, ble imidlertid sterkt oppregulert I LR under alle fem utviklingsstadier: 6.11, 4.69, 7.47, 9.05 og 8.95 (s1–s5 henholdsvis).
Metabolome-pathways
Vi gjennomførte også en komparativ interspesifikk trinnvis analyse av metabolske veier (Fig. 10). I det første utviklingsstadiet (S1) identifiserte vi 39 differensielt regulerte metabolitter. Blant de 20 beste veiene disse metabolittene var assosiert med, var flere av dem assosiert med aminosyrer, men totalt antall metabolitter per vei var relativt små (1-2), Og P-verdier antydet ikke et høyt nivå av signifikans (Fig. 10-panel 1). Spesielt vitamin B6 metabolisme (m) og mikrobiell m i ulike miljøer viste begge relativt høye p-verdier, anrikningsfaktor (EF) på 1,0 og 2 identifiserte metabolitter. I S2 identifiserte vi 58 differensielt regulerte metabolitter, assosiert med bare fire veier: tryptofan m, fenylpropanoidbiosyntese (b), b av fenylpropanoider( disse er to forskjellige veier I kegg-databasen)og fenylalanin, tyrosin og tryptofan b (ALLE EF = 1,0, 2-3 metabolitter og p < 0,5; Fig. 10-panel 2). I S3 identifiserte vi 59 differensielt regulerte metabolitter, assosiert med 19 veier, mest alle MED EF 1,0, men relativt ikke-signifikante P-verdier (> 0,5; Fig. 10-panel 3). Veier med relativt høyt antall metabolitter (n = 5) var: proteinfordøyelse og absorpsjon, b av sekundære plantemetabolitter, b av antibiotika og b av aminosyrer. I S4 identifiserte vi 58 differensielt regulerte metabolitter, assosiert med et stort antall veier, for DET meste MED EF 1.0, og relativt høye signifikansverdier (for det meste P > 0.5; Fig. 10-panel 4). Veier med relativt høyt antall metabolitter (n ≥ 3) var: fenylpropanoid b, fenylalanin, tyrosin og tryptofan b, glukosinat b, b av alkaloider avledet fra shikimatvei og 2-oksokarboksylsyre m. i moden frukt (S5) identifiserte vi 39 differensielt regulerte metabolitter, assosiert med et stort antall veier, men for det meste med lave p-verdier og bare 1 metabolitt per vei (Fig. 10-panel 5). Veier med mer enn 1 metabolitt var: proteinfordøyelse og absorpsjon, fenylpropanoid b, mineralabsorpsjon, sentral karbon m i kreft, b av sekundære metabolitter, b av fenylpropanoider og aminoat tRNA b. hovedkomponentanalysen (pca) av alle data (2 arter × 5 stadier × 5 biologiske replikater) viste høy likhet blant biologiske replikater (klynging) og bekreftet bemerkelsesverdig variasjon mellom forskjellige fruktmodningstrinn for begge arter (Fig. 10-panel 6).

Komparativ analyse AV kegg metabolisk vei berikelse. De 15 beste banene beriket Med L. ruthenicum i forhold Til l. barbarum er vist til venstre (rød), og de som er beriket Med L. barbarum i forhold Til l. ruthenicum til høyre (grønn). Utviklingsstadier (1-5) er angitt i figuren. q-verdi er EN fdr-justert p-verdi.
Metabolom-individuelle metabolitter
listen (Supplerende Datasett S6) av de mest differensielt regulerte metabolittene mellom de to artene viste en viss variasjon mellom de fem fruktutviklingsstadiene. Interessant Nok Var Fruktose 1-fosfat den mest oppregulerte metabolitten i LR, sammenlignet med LB, i alle fem stadier: log2-Foldsendring = 6,3, 7,6, 7,7, 8,1 og 6,5 (henholdsvis trinn 1 til 5). Når det gjelder metabolittene som er sterkt oppregulert i LB, var det mer variasjon mellom stadier: i S1 var forskjellene ganske små, med 9-Decenol som den mest oppregulerte metabolitten(log2 Fold change = 2.7; sammenlignet MED LR). Analyser av S2 og S3 ga svært kongruente resultater, med fenol (henholdsvis 3,7 og 3,2) som den mest oppregulerte metabolitten. I S4 var indoksylsulfat (4,7) den øverste metabolitten oppregulert i LB. Et metabolsk skifte ble observert i moden frukt (s5), hvor listen over metabolitter oppregulert i LB ble toppet av stearoylkarnitin (7,1), Metoksyeddiksyre (5,3), S-Metyl-5′-tioadenosin (4,7), lisinopril (4.7), Adenosin 3′, 5 ‘ – syklisk fosfat (cAMP) (4.7), etc. Andre metabolitter sterkt oppregulert I LR (bortsett Fra Fruktose 1-fosfat) var naringin (6,2), lauroyl-CoA (4,8), L-Phneylalanin (4,6), etc.