Figur 4: ögats struktur.Bilden är tagen från Chirstie (ref. 4) och är copyrightrestricted enligt källan (dvs. det är inte författarnas eget arbete).
ögats struktur: området i ögat (se figur 4) som upptäcker ljus och färg kallas näthinnan. De två typerna av detektionscell närvarande, stavar och kottar, bearbetar information som kommer genom linsen och skickar den ner i optisk nerv till hjärnan. Stavceller (avsom det finns cirka 100 miljoner) upptäcker graden av ljushet som kommer in iögat och deras känslighet är beroende av mängden Rhodopsin närvarande som genereras i cellerna. Rhodopsin förstörs emellertid avblekning vid exponering för ljus och därför fungerar stavceller endast i svagt ljus somvid hög belysning leder den reducerade nivån av detta ljuskänsliga pigment till avery låg känslighet. Konceller (av vilka det finns cirka 3 miljoner) är också känsliga för ljusnivåer men behåller sin funktion upp till hög belysning via användning av pigmentjodopsin.Detektering av färg är en funktion av de tre typerna av konceller som finns inom näthinnan: mellan dem täcker de det synliga spektrumet. Detta beror på att varje typ är känslig för ett annat våglängdsområde med maxmotsvarande rött (långt), grönt (medium) eller blått (kort).
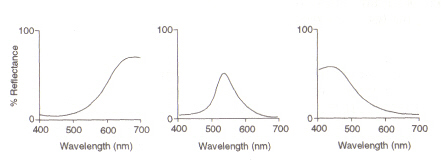
Figur 5: maximum av (frånvänster) röda, gröna och blå konceller, respektive. Bilden är tagen från Chirstie (ref. 4) och är upphovsrättsbegränsad enligt den angivna källan (dvs. det är inte författarnas eget arbete).
mekanismen för Rhodopsin i Rodcells: proteinet Rhodopsin innehåller protonatedretinal-Schiffs baskomplex som naturligt ligger i intermembranfickan bildad av de sju trans-membran a-helicalreceptorerna. Det finns många platta skivor av rhodopsin inom det yttre segmentet av arod-cell som vid ljusdetektering genomgår en foto-isomer förändring från Rhodopsin (11-cis) till all-trans retinal. Efter fotoisomeriseringskaskaden som inträffar via 5 kortlivade intermediater (flödesschema 1) diffunderar trans retinal bort och omvandlas tillbaka till 11-cisretinal innan återinträde i cykeln. Denna process sker via reduktion tillall-trans retinol följt av oxidation/isomerisering i mörkret. PhotoexcitedRhodopsin (4: e av de 5 intermediärerna) utlöser en enzymatisk kaskadprocess som resulterar i hydrolys av GMP. Detta stänger i sin tur katjonspecifika kanaler inom stavcellmembranet som är naturligt öppna för tillströmning av Na+i mörkret, och på grund av effekten av hyperpolarisering skickar den inre synatiska kroppen en nervsignal till andra neuroner i näthinnan. Slutligen hjälper den ljusinducerade minskningen av kalciumnivåer återhämtning av upphetsade neuroner till ett passivt, ”mörkt”tillstånd och cykeln startar igen vid detektering av ljus. Defotoreceptorer av konceller är också sju a-helicalreceptorer med 11-cis-retinal som deras kromofor. Detekteringsområdet varierarfrån grönt till rött eftersom de tre icke-polära hydroxylinnehållande resterna nearretinal ersätts sekventiellt med polära.

vänster-Figur 6: Position av 11-cis retinal inom Rhodopsin. Denna bild är tagen från Stryer (ref. 6) och är upphovsrättsbegränsad enligt den angivna källan (dvs. det är inte författarnas eget arbete).
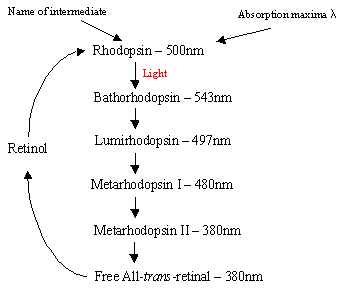
Höger-Flödesschema1: De 5 mellanprodukterna i Rhodopsincykeln.
var kommer Rhodopsin från?
människor kan inte makeRhodopsin, istället använder de och extern källa, b-karoten, som finns i mat för att syntetisera det:
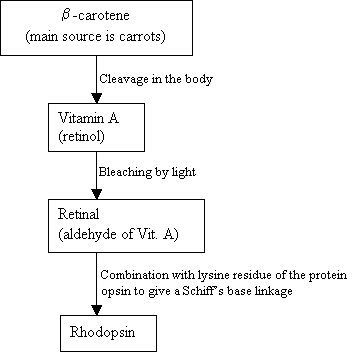 |
flödesschema 2: syntes av Rhodopsin ny bild =Figur 7: strukturerna av b-karoten (topp), VitaminA (mitten) och 11-cis retinal (botten). |
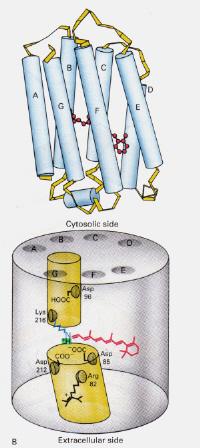
hur skiljer sig mekanismen avbakterierhodopsin som finns i Halobakterier från Rhodopsin som finns irodceller i det mänskliga ögat?: Den här gången blockerar den protonerade retinal-Schiffs baskomplex naturligt en kanal genom membranet som annars bildas av två angränsande kamrar. Det protonerade trans-komplexet donerar en proton till asp-85 som sedan tillåter utgång av samma proton till den extracellulära sidan. Fotoisomerisering till 13-cis-strukturen gör det möjligt för Schiffs bas att plocka upa proton från asp-96-återstoden på den cytosoliska sidan. Vid omorientering av cis-formen till trans fortsätter cykeln av isomerisering och protonpumpning.