vi samlade frukter av L. barbarum och L. ruthenicum i fem utvecklingsstadier, från ung frukt (10 dagar efter blomning) till mogen (mogen) frukt (34-45 dagar efter blomning) och studerade deras transkriptom och metabolom.
- RNA-seq de novo montering och funktionell annotering av unigenes
- Lycium barbarum( LB): parvisa interstegsanalyser
- LB-transkriptom
- LB Metabolome
- Lycium ruthenicum (LR): parvisa interstegsanalyser
- LR-transkriptom
- LR Metabolome
- interspecifik jämförande analys av DEGs vid olika utvecklingsstadier
- totala DEGs under fruktutvecklingen
- Transkriptomvägar
- transkriptom-enskilda gener
- Metabolome-vägar
- Metabolome-individuella metaboliter
RNA-seq de novo montering och funktionell annotering av unigenes
vi förberedde totalt 30 cDNA-bibliotek från frukter av L. barbarum och L. ruthenicum, med tre biologiska replikat (tre frukter från tre träd) vid varje tidpunkt: 2-arter 2-tidpunkter 3-biologiska replikat. Prover märktes LB/LR(1-5)-(1-3), där LB är L. barbarum och LR är L. ruthenicum, 1-5 är utvecklingsstadier av frukt (S1–S5) och 1-3 är enskilda prover (biologiska replikat); så till exempel representerar LB1-1 L. barbarum, 1: A samplade utvecklingsstadiet (S1), fruktprov nr 1 (av tre). Vi genererade över 1.72 miljarder par-end läser för dessa 30 cDNA-bibliotek, vilket motsvarar i genomsnitt 57.2 miljoner läser per prov (kompletterande Dataset S1). Sträng kvalitetsbedömning och datafiltrering gav totalt 801,766 högkvalitativa läsningar med medellängden 730 och N50 av 1107 bp (Tabell 1). Slutligen erhölls totalt 326,276 unigenes med den genomsnittliga längden på 596 bp och N50 av 847 bp från transkripten (Tabell 1). Korrelationskoefficienter för RNA-seq-data för de 30 proverna indikerar mycket god konsistens av resultat bland biologiska replikat (Fig. 2).

värmekarta av korrelationskoefficienter för RNA-seq-data för 30 prover av L. barbarum (LB) och L. ruthenicum (LR) frukter vid fem olika utvecklingsstadier. Proverna är märkta LB / r1–5_1-3, där LB är L. barbarum, LR är L. ruthenicum, 1-5 är fruktutvecklingsstadier och 1-3 individuella prover. Prover grupperades efter hierarkisk gruppering; dendrogram ovan och vänster om värmekartan indikerar släktskap av prover.
bland alla 326 276 unigenes som frågades mot offentliga databaser matchade totalt 193 021 (59,15%) gener och/eller proteiner i minst en databas och 12 171 (3,73%) kommenterades i alla databaser. Det största antalet unigenes (149 863, 45,93%) antecknades i NT-databasen och det lägsta antalet (24 017; 7,36%) i KOG-databasen.
Lycium barbarum( LB): parvisa interstegsanalyser
LB-transkriptom
det högsta antalet DEGs identifierades i alla parvisa jämförelser av 1: a etappen och i 2: A mot 5: e stegsjämförelsen (alla > 10 000 DEGs; Tabell 2). De minsta siffrorna identifierades i jämförelser mellan 3: e och 4: e och 5: e etappen (255-257). Heatmap-analys av DEGs i LB visar att ganska olika uppsättningar gener var högt uppreglerade i de tidiga utvecklingsstadierna (1 + 2) och i senare stadier (3 till 5) (Fig. 3A). Provrelaterad analys indikerar att proverna kan delas in i två klader (steg 1 + 2 och 3 + 4 + 5), med den senare kladen ytterligare indelad i två klader: steg 3 + 4 och steg 5. Intraspecifik Kegg funktionell klassificeringsanalys av dessa DEGs identifierade 15 vägar signifikant (P < 0,05) differentiellt reglerad mellan olika utvecklingsstadier (kompletterande Dataset S2). Särskilt starkt differentiellt reglerade var ’växthormonsignaltransduktion’, ’fenylpropanoidbiosyntes (b.)’, ’linolsyrametabolism (m.)’, ’ stärkelse och sackaros m.’, och ’zeatin b.’ (Fig. 3A).

värmekartor och funktionella väganalyser av differentiellt uttryckta gener (DEGs) i Lycium barbarum (panel A) och L. ruthenicum (panel B) frukter. Värmekartor genererades av en hierarkisk analys av DEGs (y-axel) och individuella prover (x-axel), där dendrogram ovan och vänster om värmekartan indikerar samspelets släktskap. Proverna är märkta LB / r_1–5_1-3, där artens akronym (LB eller LR) följs av fruktutvecklingsstadiet (1-5) och provnumret (1-3). Intraspecifika Kegg-väganalyser av DEGs i alla fem utvecklingsstadierna i de två arterna visas till höger om värmekartorna. Endast de 15 bästa berikade vägarna listas. q-värdet är ett FDR-justerat p-värde.
för en mer djupgående analys av data fokuserade vi på jämförelsen av de mest väsentligt reglerade vägarna i successiva utvecklingsstadier. I den första parvisa jämförelsen (S1 vs S2) var ’fenylpropanoid b.’ den mest differentiellt reglerade vägen, följt av ’stärkelse och sackaros m.’ (Fig. 4). Mycket stort antal DEGs (>100) identifierades i båda vägarna. Ett liknande resultat observerades i följande parvis jämförelse, S2 vs. S3, men trots det ganska stora antalet DEGs (>80) ’stärkelse och sackaros m.’uppvisade något lägre q-värde. I jämförelsen i S3 vs. S4 var’ kolfixering i fotosyntetiska organismer’ den mest signifikant reglerade vägen, men antalet gener var mycket lägre. I det sista paret var S4 vs. S5, ’zeatin b.’, flavonoid b.’, fettsyra b.’ och ’galaktos m.’ de mest signifikant reglerade vägarna, men ingen av vägarna uppvisade mer än två grader.

LB transkriptom: parvis interstegs Kegg metabolisk väg anrikning. (A) Steg 1 vs. steg 2. (B) steg 2 vs. steg 3. (C) steg 3 vs. steg 4. (D) steg 4 vs. steg 5. Topp 15 (eller alla om totalt <15) signifikant berikade vägar visas. Färgen på stapeln representerar storleken på q-värdet (ett FDR-justerat p-värde). Färgkarta visas i figuren.
LB Metabolome
vi genomförde parvisa utvecklingsstadiejämförelser för att identifiera anrikningen av metaboliter mellan alla scenpar. Antalet differentiellt reglerade metaboliter i parvisa jämförelser av utvecklingsstadier i LB varierade från 66 (steg 3 mot 4) till 129 (steg 2 mot 5). Det största antalet i successiva stegjämförelser observerades mellan 1: A och 2: a steget (117). Dessa tilldelades ett stort antal vägar; med det största antalet metaboliter tilldelade ’ B. av sekundära metaboliter ’(även det lägsta p-värdet), följt av ’protein matsmältning och absorption’ och ’B. av aminosyror’ (Fig. 5; Kompletterande Siffror). I det successiva steget par jämförelse (S2 vs. S3), ’ABC transportörer’ var den mest signifikant berikad väg, följt av’purin m.’. I S3 vs. S4-parjämförelsen var ’mikrobiell m. i olika miljöer’ den mest signifikant berikade vägen, följt av’carbapenem m.’. I S4 vs. S5-parjämförelsen uppvisade ett relativt stort antal vägar liknande resultat (två metaboliter och liknande p-värden), men anmärkningsvärt är utseendet av ’isoflavonoid b.’ och ’flavonoid b.’ bland dem (Fig. 5; Kompletterande Siffror).

LB metabolomiska data: parvis interstage Kegg metabolisk väg anrikning. (A) Steg 1 vs. steg 2. (B) steg 2 vs. steg 3. (C) steg 3 vs. steg 4. (D) steg 4 vs. steg 5. Rik faktor är förhållandet mellan antalet signifikant reglerade metaboliter i vägen och det totala antalet metaboliter som kommenteras i den vägen (intervall = 0 till 1,0). Punktens storlek representerar antalet signifikant berikade metaboliter i motsvarande väg, och punktens färg representerar P-värdet (båda legenderna visas i figuren).
när det gäller enskilda metaboliter, i jämförelsen S1 vs. S2, toppades listan över metaboliter som uppreglerades i S1 av trehalos, Glaktinol och L-äppelsyra (alla 20 log2fc-produkter i 20-årsåldern). Oljesyra, 2-Oxoadipinsyra och stearinsyra var de mest uppreglerade metaboliterna i S2 (alla 15 till 18 log2FC-sekret). Listan över de mest uppreglerade metaboliterna i S2 jämfört med S3 toppades av dihydroxiaceton, LysoPC(18:1(9Z)) och adenin (alla 16 till 17,5 log2FC). Listan över de mest uppreglerade metaboliterna i S3 (jämfört med S2) toppades av trehalos, Galaktinol och L-äppelsyra (alla 19-20 log2fc-enheter i 20-årsåldern). I jämförelsen S3 vs. S4 toppades listan över högst uppreglerade metaboliter i S3 av L-äppelsyra, DL-arginin och oljesyra (alla 18 till 19 log2FC-moduler i 19-2), medan listan över högst uppreglerade metaboliter i S4 toppades av D-mannos, N-Acetyllaktosamin och LysoPC(18:1(9Z)) i LB (alla 16 log2fc-moduler i 16). I S4 vs. S5 jämförelse, listan över de mest uppreglerade metaboliterna i S4 toppades av trehalos, palmitinsyra, N-Acetyllaktosamin (alla 17 till 18 log2FC i lb. Listan över de mest uppreglerade metaboliterna i S5 toppades av L-Norleucin, Antranilsyra (Vitamin L1) och DL-arginin (alla 16 till 18 log2FC) (kompletterande Dataset S3).
Lycium ruthenicum (LR): parvisa interstegsanalyser
LR-transkriptom
Heatmap-analys av DEGs i LR visar att nästan helt olika uppsättningar gener var mycket uppreglerade i de tidiga utvecklingsstadierna (1 + 2) och i mogen frukt (steg 5), med en uppenbar transkriptomisk återställning som inträffade efter det andra steget (Fig. 3B). Provrelaterad analys indikerar att proverna kan delas in i två klader (steg 1 + 2 och 3 + 4 + 5), med den senare kladen ytterligare indelad i två klader: steg 3 + 4 och steg 5. Intraspecifik Kegg funktionell klassificeringsanalys av dessa DEGs identifierade 35 vägar signifikant (P < 0.05) differentiellt reglerad mellan olika utvecklingsstadier (kompletterande Dataset S2). Särskilt starkt differentiellt reglerad var ’fotosyntes’, ’linolsyra m.’, ’isokinolinalkaloid b.’, ’flavonoid b.’ och ’stärkelse och sackaros m’ (Fig. 3B). Det högsta antalet DEGs identifierades i de flesta parvisa jämförelser av steg 1 (1 mot 2 var ett undantag) och i steg 2 mot 5 jämförelse (alla >5000 DEGs; Tabell 2). Det överlägset minsta antalet identifierades i S3 vs. S4-jämförelsen (39). I S1 vs S2 parvis jämförelse var ’aminosocker och nukleotidsocker m.’ den mest differentiellt reglerade vägen, följt av ’linolsyra m.’ (Fig. 6). I S2 vs. S3 parvis jämförelse var de mest signifikant differentiellt reglerade vägarna ’isokinolinalkaloid b.’,’ tyrosin m.’,’ fenylpropanoid b ’och’flavonoid b.’. I båda parvisa jämförelserna identifierades det högsta antalet DEGs (>15 respektive >35) i ’stärkelse och sackaros m.’. I S3 vs. S4 jämförelse, ’(alfa -) linolsyra m. ’ var den mest signifikant reglerade vägen, men antalet gener var mycket lägre. I jämförelsen S4 vs. S5 var’ fenylpropanoid b ’(även det största antalet DEGs) och’ linolsyra m. ’ de mest signifikant reglerade vägarna.
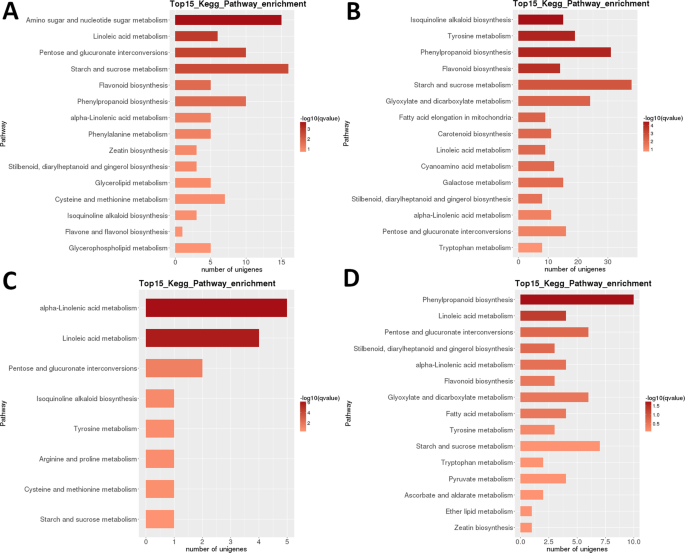
LR transkriptom: parvis interstegs Kegg metabolisk väg anrikning. (A) Steg 1 vs. steg 2. (B) steg 2 vs. steg 3. (C) steg 3 vs. steg 4. (D) steg 4 vs. steg 5. Topp 15 (eller alla om totalt <15) signifikant berikade vägar visas. Färgen på stapeln representerar storleken på q-värdet (ett FDR-justerat p-värde, färgdiagram som visas i figuren).
LR Metabolome
antalet differentiellt reglerade metaboliter i parvisa jämförelser av utvecklingsstadier i LR varierade från 66 (steg 3 mot 4) till 133 (steg 2 mot 5) (kompletterande Dataset S3). I successiva stegjämförelser observerades det största antalet mellan 1: A och 2: a steget (117). Dessa tilldelades ett stort antal vägar; med det största antalet metaboliter tilldelade ’B av sekundära metaboliter’, följt av’ protein matsmältning och absorption’,’ B. av aminosyror ’och’flavonoid b’. I jämförelsen S2 vs. S3 var ’ABC-transportörer ’den mest väsentligt berikade vägen, följt av’purine m’. I S3 vs. S4-parjämförelsen var ’mikrobiell m. i olika miljöer’ den mest signifikant berikade vägen, följt av’carbapenem m.’. I S4 vs. S5-par Jämförelse, ett relativt stort antal vägar uppvisade liknande resultat (2 metaboliter och liknande p-värden), men anmärkningsvärt är utseendet av ’isoflavonoid b.’ och ’flavonoid b.’ bland dem (Fig. 7; Kompletterande Siffror).

LR metabolomic data: parvis interstage Kegg metabolisk väg anrikning. (A) Steg 1 vs. steg 2. (B) steg 2 vs. steg 3. (C) steg 3 vs. steg 4. (D) steg 4 vs. steg 5. Rik faktor är förhållandet mellan antalet signifikant reglerade metaboliter i vägen och det totala antalet metaboliter som kommenteras i den vägen (intervall = 0 till 1,0). Punktens storlek representerar antalet signifikant berikade metaboliter i motsvarande väg, och punktens färg representerar P-värdet (båda legenderna visas i figuren).
när det gäller enskilda metaboliter (kompletterande dataset S3), i S1 vs. S2 jämförelse, listan över metaboliter uppreglerade i S1 toppades av trehalos, Galaktinol och L-äppelsyra (19-21 log2FC), medan oljesyra, 2-Oxoadipinsyra och stearinsyra var de mest uppreglerade metaboliterna i s2 (15-18 log2fc). I S2 vs. S3-jämförelsen var dihydroxiaceton, Indoxylsulfat och N-Acetyllaktosamin högst uppreglerade metaboliter i s2 (17-19, 5 log2fc) och trehalos, Galaktinol och L-äppelsyra (19-21 log2fc) i S3. I S3 vs. S4 jämförelse, l-äppelsyra, DL-arginin och oljesyra uppreglerades i S3 (16-19 log2fc) och 1,7-Dimetylxantin, D-mannos och N-Acetyllaktosamin (15-17 log2FC) i S4. I jämförelsen S4 vs. S5 uppreglerades Flavinmononukleotid, trehalos och Isoferulsyra i S4 (18-20 log2fc-s), medan PG(16:0/18:1(9Z)), D-prolin och DL-arginin var högst uppreglerade (alla 16-18 log2fc-s) metaboliter.
interspecifik jämförande analys av DEGs vid olika utvecklingsstadier
totala DEGs under fruktutvecklingen
interspecifik parvis stegsjämförelse (LR1 vs. LB1, LR2 vs. LB2, etc.) visar att 928 DEGs delades av alla fem paren (Fig. 8A). Det högsta antalet DEGs identifierades i steg 3 (3989) och det lägsta i steg 4 (2825) (Fig. 8B); medan det högsta antalet DEGs som är unika för ett par observerades i steg 3 (574), 1 och 5 (båda 554) och det lägsta i steg 4 (126) (Fig. 8A). Antalet upp-och nedreglerade DEGs var relativt lika i var och en av de parvisa scenjämförelserna; t.ex. i steg 5 uppreglerades 1668 DEGs och 1670 DEGs nedreglerades i LR jämfört med LB (Fig. 8B). Men i de andra fyra stadierna var antalet uppreglerade gener något (93 till 189 DEGs) högre.

gener differentiellt uttryckta (DEGs) mellan frukter av L. barbarum (LB) och L. ruthenicum (LR). (A) detaljerade stagewise (1 till 5) jämförelser (LB vs. LR). (B) antalet uppreglerade (röda) och nedreglerade (gröna) deg i LR jämfört med LB i fem studerade utvecklingsstadier.
Transkriptomvägar
värmekartan analys av DEGs indikerar att frukterna av två arter uppvisar mycket olika genuttrycksprofiler under alla utvecklingsstadier, men biologiska replikat uppvisade mycket liknande profiler, vilket indikerar en begränsad mängd individuell variation i varje utvecklingsstadium (kompletterande figurer: Fig. S9). Jämförande analys av Kegg-väganrikning visar att endast vissa vägar konsekvent var mycket berikade (i termer av genreglering) i LR jämfört med LB under alla fem utvecklingsstadier (Fig. 9). I synnerhet växthormonsignaltransduktion (2: A högsta i S1, 8: e högsta i S2, den högsta i S3, S4 och S5) och växtpatogeninteraktion (den högsta i S1, 3: e högsta i S2, 2: A högsta i S3, 4: e högsta i S4 och 15: e högsta i S5) var relativt högt uppreglerade i alla steg. Fenylpropanoidbiosyntes (inte i topp 15 i S1, den högsta i S2, 3: e högsta i S3, 7: e högsta i S4, 11: e högsta i S5), ubikinon och annan terpenoid-kinonbiosyntes (inte i topp 15 i S1, 6: e högsta i s2, 7: e högsta i S3, 2: A högsta i S4, 6: e högsta i S5) var också relativt högt uppreglerade i alla steg utom den första. Flavonoid biosyntesvägen var inte mycket berikad i tidiga stadier (inte i topp 15 i S1, 14: e högst i S2) och mycket berikad i sena steg (3: e till 4: e högst under Steg 3 till 5). (alfa-)linolsyrametabolism var mycket berikad i mellanstadier (9: e högsta i S1, 2: A högsta i S2, 5: e och 6: e högsta i S3, 8: e högsta i S4, inte i topp 15 i S5).

jämförande analys av Kegg metabolisk väg anrikning. De 15 bästa vägarna berikade i L. ruthenicum jämfört med L. barbarum visas till vänster (röd) och de berikade i L. barbarum jämfört med L. ruthenicum till höger (grön). Utvecklingsstadier (1-5) anges i figuren. q-värdet är ett FDR-justerat p-värde.
bland vägarna nedreglerade i L. ruthenicum i jämförelse med L. barbarum (Fig. 9) anmärkningsvärda förändringar observerades mellan de tidiga stadierna (1 och 2), när cyanoaminosyrametabolism och karotenoidbiosyntes var mest nedreglerade och sena steg (4 och 5), när VIRVELINTERAKTIONER i vesikulär transport, nikotinat och nikotinamidmetabolism och porfyrin-och klorofyllmetabolism var konsekvent relativt högt nedreglerade.
transkriptom-enskilda gener
bland de mest differentiellt uttryckta generna var vissa utvecklingsfasspecifika (dvs. mycket differentiellt reglerade endast i tidiga eller sena utvecklingsstadier), men vissa reglerades konsekvent mycket differentiellt genom alla fem studerade stadier (kompletterande Dataset S4). Flera immunitetsrelaterade gener mycket högt uppreglerade i LR jämfört med LB i de tidiga utvecklingsstadierna är bland exemplen på det utvecklingsstadiespecifika uttrycksmönstret: chitinase var den 2: A högsta uppreglerade DEG i S1 (13,43-faldigt), den högsta i S2 (13,89-faldigt), men i senare skeden var det inte en DEG. På samma sätt var EIX-receptorn 1/2 också mycket högt uppreglerad i tidiga stadier, Den högsta i S1 (13.70) och 2nd-högst i S2 (10.90), men det identifierades inte heller som DEG i senare stadier. Vissa tillväxtrelaterade gener uppvisade också ett liknande uttrycksmönster: Fosfoglyceratkinas (PGK) var bland de handfulla mest uppreglerade generna i de tre första stegen (13.14, 12.87 och 12.77 respektive), men det identifierades inte heller som DEG i senare skeden. CCR4-inte transkriptionskomplex subenhet 7/8 (CNOT7/8) uppvisade också ett mycket liknande uttrycksmönster: högt uppreglerat i tre första steg och inte en DEG i steg 4 och 5. Flera flavonoid-och fenylpropanoidbiosyntesassocierade gener uppvisade ett omvänd utvecklingsstadiespecifikt uttrycksmönster, med relativt lågt uttryck i tidiga stadier och mycket högt i senare stadier. Exempel är: bifunktionell dihydroflavonol 4-reduktas / flavanon 4-reduktas (DFR), som uppreglerades något i LR i S1 (2.25), inte en DEG i S2, högt uppreglerad i S3 (7.79) och 3: e högsta uppreglerade genen i S4 (14.25) och S5 (16.03). En paralog av denna gen uppvisade ett nästan identiskt mönster: något uppreglerat i S1 (2.44), inte en DEG i S2, högt uppreglerad i S3 (7.40), 6th-högst uppreglerad DEG i S4 (13.26) och 5th i S5 (14.59). På samma sätt var flavonoid 3′,5′-hydroxylas (F3’5′ h) inte en DEG i de två första stegen, högt uppreglerad i S3 (6.69), 5th-högst uppreglerad gen i S4 (13.42) och 4th-högst i S5 (15.05). Flavonoid O-metyltransferas (OMT) var inte en DEG i S1, men i S2 uppvisade den redan en medelhög uppregleringsnivå (4.32), av S3 var den redan den tredje högsta uppreglerade DEG (13.30), och det var den högst uppreglerade genen i S4 (18.73) och S5 (18.10). Leucoanthocyanidindioxygenas (ldox; antocyaninbiosyntes) var inte en DEG i S1 och S2, följt av hög till mycket hög uppreglering i senare skeden (5,63, 9,44, 11,56, respektive). Två kalkonsyntasparaloger (CHS och CHS2; flavonoidbiosyntes) var inte heller starkt reglerade i S1 och S2 (CHS2: inte en DEG, CHS: -1.14 i S1, inte en DEG i S2), men i S3–S5 uppvisade båda generna en medelhög till hög uppreglering (CHS2: 5.32, 7.84, 6.00; och CHS: 4.67, 7.01, 6.82; respektive). Vi valde dessa gener för qPCR-analys, och resultaten är mycket kongruenta med RNA-seq-data (kompletterande resultat; kompletterande Dataset S5). Slutligen uppreglerades också cytokinindehydrogenas, en zeatinbiosyntesrelaterad gen, alltmer under de tre sista stegen (2,6-5,7).
vissa gener uttrycktes emellertid konsekvent differentiellt under alla fem studerade stadier. Exempel inkluderade också några immunitetsrelaterade gener, såsom två paraloger av glutation S-transferas, högt uppreglerade i LR jämfört med LB i alla steg: 9.38 och 8.58 (alla värden presenterade som vikförändringar i respektive ordning) i S1, 6.30 och 6.34 i S2, den 2: A och 7: e högsta uppreglerade DEGs i S3 (14.08 och 12.70), 2: A och 4: e högsta i S4 (15.71 och 14.16) och 2: A och 6: e högsta i S5 (16.48 och 14.40). Växtsjukdomsresistensprotein RPM1 var också högt uppreglerat i alla fem steg (S1 = 13.15; S2 = 12.08; S3 = 13.11, S4 = 12.81; S5 = 13:94). Bland de konsekvent differentiellt uttryckta generna i alla utvecklingsstadier var också några relaterade till aminosyrametabolismen, men deras mönster vändes i jämförelse med tidigare exempel: de uppvisade hög nedreglering i LR jämfört med LB. Exempel är acetyl-CoA acyltransferas 1 (AAT1; valin, leucin och isoleucin nedbrytning), med en temporal profil av allt högre nedreglering, med början från -7,0 i S1 till <−10-faldigt i de tre sista stegen. Proliniminopeptidas, associerat med arginin och prolinmetabolism, var starkt nedreglerat i LR i alla steg: S1 = -9,75, S2 = -10,89 (3: e högsta), S3 = -11,05 (4: e högsta), S4 = -10,01 och S5 = -11,98 (3: e högsta). Slutligen var 5–metyltetrahydropteroyltriglutamat-homocysteinmetyltransferas (metE) konsekvent extremt högt nedreglerad i LR i alla steg: 2: A högsta i S1 (-11.76), den högsta i S2 (-11.74), 3: e högsta i S3 (-11.43), den högsta i S4 (-12.36) och 2: A högsta i S5 (-12.83). Två DNA-replikation och transkriptionsassocierade gener var också starkt nedreglerade i LR i alla steg: GTP-bindande kärnprotein Ran (RAN; -10.0 till -12.0) och replikationsfaktor A1 (RFA1) (-8.0 till -12.0). Vissa tillväxt-och stressrelaterade gener var också konsekvent starkt nedreglerade i LR: heterogent kärnribonukleoprotein A1/A3 (hnRNP; -7 till -11) och värmechock 70 kDa-protein 1/8 (HSPA1_8) S1 = -4,95, S2 = -8,88, S3 = -11,48 (2: A högsta), S4 = -9,36, S5 = -12,89 (den högsta). Intriguingly var en fenylpropanoid biosyntesrelaterad gen, shikimate hydroxycinnamoyltransferas (HCT), också konsekvent starkt nedreglerad i LR: S1 = -6.82, S2 = -8.14, S3 = -11.71 (den högsta), S4 = -11.00 (3rd-highest), S5 = -11.91 (4th-highest). En nyckelregulator för antocyaninbiosyntes, transkriptionsfaktor MYB114, var emellertid högt uppreglerad i LR under alla fem utvecklingsstadier: 6.11, 4.69, 7.47, 9.05 och 8.95 (S1–S5 respektive).
Metabolome-vägar
vi genomförde också en jämförande interspecifik stegvis analys av metaboliska vägar (Fig. 10). I det första utvecklingsstadiet (S1) identifierade vi 39 differentiellt reglerade metaboliter. Bland de 20 bästa vägarna var dessa metaboliter associerade med, flera av dem var associerade med aminosyror, men det totala antalet metaboliter per väg var relativt små (1-2) och P-värden antydde inte en hög nivå av betydelse (Fig. 10-panel 1). I synnerhet vitamin B6-metabolism (m) och mikrobiell m i olika miljöer uppvisade båda jämförelsevis höga P-värden, anrikningsfaktor (EF) på 1,0 och 2 identifierade metaboliter. I S2 identifierade vi 58 differentiellt reglerade metaboliter, associerade med endast fyra vägar: tryptofan m, fenylpropanoidbiosyntes (b), B av fenylpropanoider (dessa är två olika vägar i KEGG-databasen) och fenylalanin, tyrosin och tryptofan b (Alla EF = 1,0, 2-3 metaboliter och p < 0,5; Fig. 10-panel 2). I S3 identifierade vi 59 differentiellt reglerade metaboliter, associerade med 19 vägar, mest alla med EF 1.0, men relativt icke-signifikanta P-värden (>0.5; Fig. 10-panel 3). Vägar med relativt högt antal metaboliter (n = 5) var: protein matsmältning och absorption, b av växt sekundära metaboliter, B av antibiotika och B av aminosyror. I S4 identifierade vi 58 differentiellt reglerade metaboliter, associerade med ett stort antal vägar, mestadels med EF 1.0 och jämförelsevis höga signifikansvärden (mestadels P > 0.5; Fig. 10-panel 4). Vägar med relativt högt antal metaboliter (n 2BG) var: fenylpropanoid b, fenylalanin, tyrosin och tryptofan b, glukosinat b, B alkaloider härledda från shikimatvägen och 2-oxokarboxylsyra m. i den mogna frukten (S5) identifierade vi 39 differentiellt reglerade metaboliter, associerade med ett stort antal vägar, men mestadels med låga P-värden och endast 1 metabolit per väg (Fig. 10-panel 5). Vägar med mer än 1 metabolit var: proteinförtunning och absorption, fenylpropanoid b, mineralabsorption, centralt kol m i cancer, b av sekundära metaboliter, B av fenylpropanoider och aminoacy tRNA b. huvudkomponentanalysen (PCA) av alla data (2-arter i 5-steg i 5-biologiska replikat) avslöjade hög likhet mellan biologiska replikat (kluster) och bekräftade anmärkningsvärd variation mellan olika fruktmognadssteg för båda arterna (Fig. 10-panel 6).

jämförande analys av Kegg metabolisk väg anrikning. De 15 bästa vägarna berikade i L. ruthenicum jämfört med L. barbarum visas till vänster (röd) och de berikade i L. barbarum jämfört med L. ruthenicum till höger (grön). Utvecklingsstadier (1-5) anges i figuren. q-värdet är ett FDR-justerat p-värde.
Metabolome-individuella metaboliter
listan (kompletterande Dataset S6) av mest differentiellt reglerade metaboliter mellan de två arterna, uppvisade viss variation bland de fem fruktutvecklingsstadierna. Intressant var fruktos 1-fosfat den mest uppreglerade metaboliten i LR, jämfört med LB, under alla fem steg: log2 Fold change = 6.3, 7.6, 7.7, 8.1 och 6.5 (steg 1 till 5 respektive). När det gäller de metaboliter som är mycket uppreglerade i LB var det mer variation mellan steg: i S1 var skillnaderna ganska små, med 9-Decenol som den mest uppreglerade metaboliten (log2-faldig förändring = 2.7; jämfört med LR). Analyser av S2 och S3 gav mycket kongruenta resultat, med fenol (3,7 respektive 3,2) som den mest uppreglerade metaboliten. I S4 var indoxylsulfat (4,7) den toppmetabolit som uppreglerades i LB. En metabolisk förändring observerades i den mogna frukten (S5), där listan över metaboliter som uppreglerades i LB toppades av stearoylkarnitin (7.1), Metoxiättiksyra (5.3), S-metyl-5′-tioadenosin (4.7), lisinopril (4.7), adenosin 3′, 5′-cykliskt fosfat (cAMP) (4.7), etc. Andra metaboliter högt uppreglerade i LR (bortsett från fruktos 1-fosfat) var naringin (6.2), lauroyl-CoA (4.8), L-Phneylalanin (4.6), etc.