shromážděné plody L. barbarum a L. ruthenicum na pět vývojových stádií, od mladých ovoce (≈10 dní po odkvětu) zralý (zralá) ovoce (34-45 dní po odkvětu), a studoval jejich transcriptome a metabolome.
- RNA-seq de novo sestavy a funkční anotace unigenes
- Lycium barbarum (LB): pairwise interstage analýzy
- LB Transcriptome
- LB Metabolome
- Lycium ruthenicum (LR): párové interstage analýzy
- LR Transcriptome
- LR Metabolome
- Mezidruhové srovnávací analýza DEGs v různých vývojových stádiích
- Celkem DEGs během vývoje ovoce
- Transcriptome – cesty
- Transcriptome – jednotlivé geny
- metabolické dráhy
- Metabolome – jednotlivé metabolity
RNA-seq de novo sestavy a funkční anotace unigenes
Jsme pro vás připravili celkem 30 cDNA knihovny z plody L. barbarum a L. ruthenicum, s tří biologických replikátů (tři ovoce ze třech stromů), v každém bodě: 2 druh × 5 čas-bodů × 3 biologických replikátů. Vzorky byly označeny LB/LR(1-5)-(1-3), kde LB je L. barbarum a LR je L. ruthenicum, 1-5 jsou vývojová stadia ovoce (S1–S5), a 1-3 jsou jednotlivé vzorky (biologické opakování); tak například LB1-1 představuje L. barbarum, 1. vzorkované vývojové stádium (S1), vzorek ovoce č. 1 (ze tří). Pro tyto knihovny cDNA 30 jsme vygenerovali více než 1,72 miliardy párových čtení, což odpovídá průměru 57,2 milionu čtení na vzorek (Doplňkový datový soubor S1). Přísné hodnocení kvality a filtrování dat přineslo celkem 801 766 vysoce kvalitních čtení s průměrnou délkou 730 a N50 1107 bp (Tabulka 1). Nakonec bylo z přepisů získáno celkem 326 276 unigenů s průměrnou délkou 596 bp a N50 847 bp (Tabulka 1). Korelační koeficienty pro data RNA – seq pro 30 vzorků naznačují velmi dobrou konzistenci výsledků mezi biologickými replikáty (obr. 2).

Heatmap korelační koeficienty pro RNA-seq dat pro 30 vzorků z L. barbarum (LB) a L. ruthenicum (LR) ovoce na pět různých vývojových fázích. Vzorky jsou označeny LB / R1-5_1-3, kde LB je L. barbarum, LR je L. ruthenicum, 1-5 jsou vývojové fáze ovoce a 1-3 jednotlivé vzorky. Vzorky byly seskupeny hierarchickým shlukováním; dendrogramy nad a vlevo od tepelné mapy naznačují příbuznost vzorků.
Mezi všemi 326,276 unigenes dotazovány proti veřejných databází, celkem 193,021 (59.15%) uzavřeno genů a/nebo proteinů v alespoň jedné databáze, a 12,171 (3.73%) byly komentovaný ve všech databázích. Největší počet unigenů (149,863, 45,93%) byl anotován v databázi NT a nejnižší počet (24,017; 7,36%) v databázi KOG.
Lycium barbarum (LB): pairwise interstage analýzy
LB Transcriptome
nejvyšší počty DEGs byly identifikovány ve všech párové srovnání 1. fázi a ve 2. vs. 5. etapa porovnání (> 10,000 DEGs; Tabulka 2). Nejmenší čísla byla zjištěna ve srovnání 3. vs. 4. a 4. vs. 5. fáze (255-257). Heatmap analýza DEGs v LB ukazuje, že poměrně odlišné sady genů byly vysoce upregulated v raných vývojových stádiích (1 + 2) a v pozdějších fázích (3 až 5) (Obr. 3A). Analýza příbuznosti vzorků ukazuje, že vzorky by mohly být rozděleny do dvou klad (fáze 1 + 2 a 3 + 4 + 5), přičemž druhý klad se dále dělí na dvě klady: stupně 3 + 4 a stupeň 5. Vnitrodruhových KEGG funkční klasifikace analýza těchto DEGs identifikovány 15 cest významně (P < 0.05) rozdílně regulovány mezi různé vývojové etapy (Doplňkové datové sady S2). Zvláště silně diferencovaně regulované byly „signální transdukce rostlinných hormonů“, “ biosyntéza fenylpropanoidů (b.)“, “ metabolismus kyseliny linolové (m.)“, “ škrob a sacharóza m.“a“ zeatin b. „(obr. 3A).

Heatmaps a funkční cesta analýzy rozdílně vyjádřené geny (DEGs) v Lycium barbarum (panel a) a L. ruthenicum (panel B) ovoce. Heatmaps byly generovány pomocí hierarchické analýzy DEGs (osa y) a jednotlivé vzorky (osa x), kde dendrograms nad a vlevo od heatmap uveďte příbuznosti vzorků. Vzorky jsou označeny LB / R_1–5_1-3, kde za zkratkou druhu (LB nebo LR) následuje vývojové stadium plodu (1-5) a číslo vzorku (1-3). Vnitrodruhových KEGG pathway analýzy DEGs ve všech pěti vývojových fázích obou druhů jsou zobrazeny na pravé straně heatmaps. Pouze top 15 obohacené cesty jsou uvedeny. q-hodnota je p-hodnota upravená FDR.
pro podrobnější analýzu dat, zaměřili jsme se na srovnání nejvýznamnějších regulovaných cest v postupných vývojových fázích. V prvním párovém srovnání (S1 vs S2) byl „fenylpropanoid b.“ nejvíce vysoce diferencovaně regulovanou cestou, následovanou „škrobem a sacharózou m.“ (obr. 4). V obou cestách bylo zjištěno velmi velké množství deg (>100). Podobný výsledek byl pozorován v následujícím párovém srovnání, S2 vs. S3, ale navzdory poměrně velkému počtu deg (>80) “ škrobu a sacharózy m.’vykazoval poněkud nižší q-hodnota. Ve srovnání S3 vs. S4, „fixace uhlíku ve fotosyntetice, organismy“ byla nejvýrazněji regulovanou cestou, ale počty genů byly mnohem nižší. V poslední dvojici, S4 vs S5, ‚zeatin b.‘, flavonoid b.‘, mastné kyseliny b. “ a “ galaktózy m.‘ byly většinou výrazně regulované cest, ale žádná z cest vystaveny více než dvě DEGs.

LB transcriptome: pairwise interstage KEGG metabolických cest obohacení. A) fáze 1 vs. fáze 2. B) fáze 2 vs. fáze 3. C) fáze 3 vs. fáze 4. D) 4.etapa vs. 5. etapa. Top 15 (nebo všechny, pokud celkem <15) jsou zobrazeny významně obohacené cesty. Barva lišty představuje velikost hodnoty q (hodnota p upravená FDR). Barevný graf je znázorněn na obrázku.
LB Metabolome
Jsme provedli párové vývojové fázi srovnání identifikovat obohacení metabolitů mezi všemi fázi páry. Počet diferencovaně regulovaných metabolitů v párovém srovnání vývojových stadií v LB se pohyboval od 66 (stadia 3 vs 4) do 129(stadia 2 vs 5). Největší počet v po sobě jdoucích stadiích srovnání byl pozorován mezi 1.a 2. stupněm (117). Ty byly přiřazeny k velkému počtu drah; s největším počtem metabolitů přiřazeny k ‚b. sekundární metabolity‘ (i nejnižší p-hodnota), následovaná ‚trávení bílkovin a vstřebávání“ a “ b. aminokyseliny (viz.Obr. 5; Doplňkové Údaje). V následném srovnání dvojic (S2 vs. S3), „ABC transportéry“ byla nejvýrazněji obohacená cesta, následovaná „purinem m.“. V porovnání párů S3 vs. S4 byl „mikrobiální m. v různých prostředích“ nejvýrazněji obohacenou cestou, následovanou „karbapenem m.“. V S4 vs S5 pár srovnání, relativně velký počet cest vykazovaly podobné výsledky (dva metabolity a podobné p-hodnoty), ale pozoruhodný je vzhled ‚isoflavonoid b.“ a “ flavonoidů b.‘ mezi nimi (Obr. 5; Doplňkové Údaje).

LB metabolomické údaje: párové mezistupňové obohacení metabolické dráhy KEGG. A) fáze 1 vs. fáze 2. B) fáze 2 vs. fáze 3. C) fáze 3 vs. fáze 4. D) 4.etapa vs. 5. etapa. Rich faktor je poměr počtu významně regulovaných metabolitů v dráze a celkového počtu metabolitů anotovaných v této cestě (rozmezí = 0 až 1,0). Velikost bodu představuje počet významně obohacených metabolitů v odpovídající dráze a barva bodu představuje hodnotu P (obě legendy znázorněné na obrázku).
pokud jde o jednotlivé metabolity, v S1 vs. S2 srovnání, seznam metabolitů upregulated v S1 byla završena tím, že Trehalosa, Glactinol a L-Jablečné kyseliny (všechny ≈20 log2FC). Kyselina olejová, kyselina 2-Oxoadipová a kyselina stearová byly nejvíce vysoce upregulovanými metabolity v S2 (všech ≈15 až 18 log2FC). Seznam nejvíce vysoce upregulated metabolitů v S2 v porovnání s S3 byla završena Dihydroxyaceton, LysoPC(18:1(9Z)), a Adenin (všechny ≈16 na 17,5 log2FC). Seznam nejvíce vysoce upregulated metabolitů v S3 (oproti S2) byla završena tím, že Trehalosa, Galactinol a L-Jablečné kyseliny (všechny ≈19 20 log2FC). V S3 vs S4 porovnávání, seznam nejvíce vysoce upregulated metabolitů v S3 byla završena L-kyselina Jablečná, kyselina DL-Arginin a kyselina Olejová (všechny ≈18 do 19 log2FC), vzhledem k tomu, že na seznamu nejvíce vysoce upregulated metabolitů v S4 byla završena D-Mannosa, N-Acetyllactosamine a LysoPC(18:1(9Z)) v LB (všechny ≈16 log2FC). V S4 vs. S5 srovnání, seznamu nejvíce vysoce upregulated metabolitů v S4 byla završena tím, že Trehalosa, Palmitové kyseliny, N-Acetyllactosamine (všechny ≈17 do 18 log2FC) v LB. Seznam nejvíce vysoce upregulated metabolitů v S5 byla završena L-Norleucinu, Anthranilic kyselina (Vitamin L1) a DL-Arginin (všechny ≈16 do 18 log2FC) (Doplňující datové sady, S3).
Lycium ruthenicum (LR): párové interstage analýzy
LR Transcriptome
Heatmap analýza DEGs v LR ukazuje, že téměř zcela odlišné sady genů byly vysoce upregulated v raných vývojových stádiích (1 + 2) a v zralé ovoce (fáze 5), se zjevným transcriptomic reset vyskytující se po druhé fázi (Obr. 3B). Analýza příbuznosti vzorků ukazuje, že vzorky by mohly být rozděleny do dvou klad (fáze 1 + 2 a 3 + 4 + 5), přičemž druhý klad se dále dělí na dvě klady: stupně 3 + 4 a stupeň 5. Vnitrodruhových KEGG funkční klasifikace analýza těchto DEGs identifikovány 35 cest významně (P < 0.05) rozdílně regulovány mezi různé vývojové etapy (Doplňkové datové sady S2). Zvláště silně rozdílně regulovány byly fotosyntézy‘, ‚kyselina linolová m.‘, ‚isochinolin alkaloid b.‘, ‚flavonoidů b.“ a „škrob a sacharóza m‘ (Obr. 3B). Nejvyšší počty DEGs byly identifikovány ve většině párového srovnání stupně 1 (1 vs. 2 byl výjimkou) a ve fázi 2 vs. 5 porovnání (>5,000 DEGs; Tabulka 2). Zdaleka nejmenší číslo bylo identifikováno ve srovnání S3 vs. S4 (39). V párovém srovnání S1 vs S2 byl „amino cukr a nukleotidový cukr m.“ nejvíce vysoce diferencovaně regulovanou cestou, následovanou “ kyselina linolová m. „(obr. 6). V párovém srovnání S2 vs. S3 byly nejvíce významně odlišně regulovanými cestami “ izochinolinový alkaloid b.“, „tyrosin m.“, „fenylpropanoid b“ a “ flavonoid b.“. V obou párového srovnání, nejvyšší počet DEGs (>15 >35) byl identifikován ve škrobu a sacharózy m.‘. V S3 vs. Srovnání S4, „(alfa-)kyselina linolová m. “ byla nejvýznamnější regulovanou cestou, ale počty genů byly mnohem nižší. Ve srovnání S4 vs. S5 byly nejvýrazněji regulovanými cestami „fenylpropanoid b“ (také největší počet deg) a „kyselina linolová m.“.
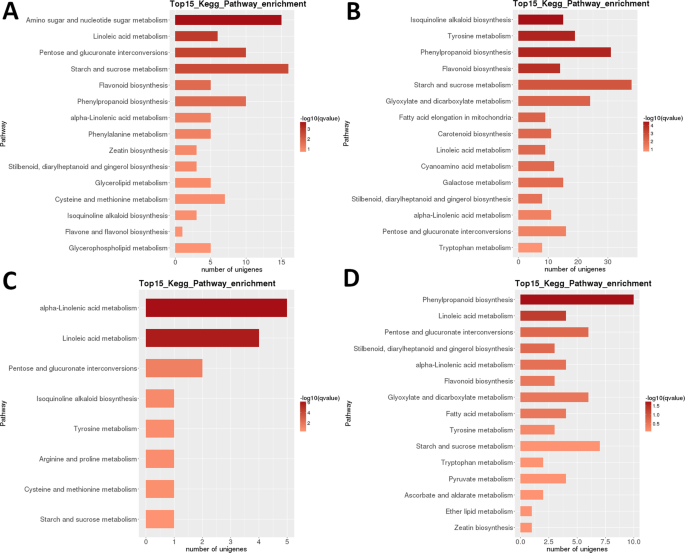
LR transcriptome: pairwise interstage KEGG metabolických cest obohacení. A) fáze 1 vs. fáze 2. B) fáze 2 vs. fáze 3. C) fáze 3 vs. fáze 4. D) 4.etapa vs. 5. etapa. Top 15 (nebo všechny, pokud celkem <15) jsou zobrazeny významně obohacené cesty. Barva lišty představuje velikost hodnoty q (hodnota p upravená FDR, barevný graf znázorněný na obrázku).
LR Metabolome
počet diferenciálně regulované metabolitů v párové srovnání vývojových etap v LR se pohybovala od 66 (fáze 3 vs 4) 133 (fáze 2 vs 5) (Doplňující datové sady, S3). V následných srovnáváních fází byl největší počet pozorován mezi 1.a 2. stupněm (117). Ty byly přiřazeny k velkému počtu cest; s největším počtem metabolitů přiřazeny k ‚b sekundární metabolity‘, následuje ‚trávení bílkovin a vstřebávání‘, ‚b. aminokyselin a flavonoidů b‘. Ve srovnání S2 vs. S3 byl „ABC transportéry“ nejvýrazněji obohacenou cestou, následovanou „purinem m“. V porovnání párů S3 vs. S4 byl „mikrobiální m. v různých prostředích“ nejvýrazněji obohacenou cestou, následovanou „karbapenem m.“. V S4 vs. S5 pár srovnání, relativně velký počet cest vykazovaly podobné výsledky (2 metabolity a podobné p-hodnoty), ale pozoruhodný je vzhled ‚isoflavonoid b.“ a “ flavonoidů b.‘ mezi nimi (Obr. 7; Doplňkové Údaje).

LR metabolomic údaje: pairwise interstage KEGG metabolických cest obohacení. A) fáze 1 vs. fáze 2. B) fáze 2 vs. fáze 3. C) fáze 3 vs. fáze 4. D) 4.etapa vs. 5. etapa. Rich faktor je poměr počtu významně regulovaných metabolitů v dráze a celkového počtu metabolitů anotovaných v této cestě (rozmezí = 0 až 1,0). Velikost bodu představuje počet významně obohacených metabolitů v odpovídající dráze a barva bodu představuje hodnotu P (obě legendy znázorněné na obrázku).
pokud jde o jednotlivé metabolity (Doplňkový datový soubor S3), v S1 vs. S2 srovnání, seznam metabolitů upregulated v S1 byla završena tím, že Trehalosa, Galactinol a L-Jablečné kyseliny (≈19-21 log2FC), vzhledem k tomu, že Olejová kyselina, 2-Oxoadipic kyseliny a Stearové kyseliny byly nejvíce vysoce upregulated metabolitů v S2 (≈15-18 log2FC). V S2 vs. S3 srovnání, Dihydroxyaceton, Indoxyl sulfát a N-Acetyllactosamine jsou většinou vysoce upregulated metabolitů v S2 (≈17-19.5 log2FC) a Trehalosa, Galactinol a L-Jablečné kyseliny (≈19-21 log2FC) v S3. V S3 vs. S4 srovnání, kyselina L-jablečná, DL-arginin a kyselina olejová byly upregulovány v S3 (≈16-19 log2FC) a 1,7-Dimethylxanthin, D-manóza a N-Acetyllaktosamin (≈15-17 log2FC) v S4. V S4 vs S5 srovnání, Flavin mononucleotide, Trehalosa a Isoferulic kyseliny jsou upregulated v S4 (≈18-20 log2FC), vzhledem k tomu, že PG(16:0/18:1(9Z)), D-Prolin, a DL-Arginin jsou většinou vysoce upregulován (všechny ≈16 do 18 log2FC) metabolitů.
Celkem DEGs během vývoje ovoce
Mezidruhové párového fázi srovnání (LR1 vs LB1, LR2 vs. LB2, atd.) ukazuje, že 928 deg bylo sdíleno všemi pěti páry (obr. 8A). Nejvyšší počet deg byl identifikován ve stupni 3 (3989) a nejnižší ve stupni 4 (2825) (obr. 8B); zatímco nejvyšší počet deg jedinečných pro pár byl pozorován ve stupních 3 (574), 1 a 5 (oba 554) a nejnižší ve stupni 4 (126) (obr. 8A). Čísla z up – a down-regulovaných DEGs byly relativně podobné v každém z párového fázi srovnání; např. ve fázi 5, 1668 DEGs byly upregulovány a 1670 DEGs byly downregulated v LR ve srovnání s LB (Obr. 8B). V ostatních čtyřech stádiích byl však počet upregulovaných genů mírně (93 až 189 deg) vyšší.

Geny rozdílně vyjádřené (DEGs) mezi plody L. barbarum (LB) a L. ruthenicum (LR). (A) podrobné fázové (1 až 5) srovnání (LB vs. LR). (B) počet upregulován (červená) a downregulated (zelená) DEGs v LR ve srovnání s LB v pěti studovaných vývojových stádiích.
Transcriptome – cesty
Heatmap analýza DEGs ukazuje, že plody dvou druhů vykazují velmi odlišné profily genové exprese v průběhu všech vývojových stádií, ale biologické opakování vykazovaly velmi podobné profily, což svědčí o omezené množství individuálních rozdílů v každé vývojové fázi (Doplňující Údaje: Obr. S9). Srovnávací analýza obohacení dráhy KEGG ukazuje, že pouze některé cesty byly trvale vysoce obohaceny (z hlediska regulace genů)v LR ve srovnání s LB ve všech pěti vývojových stádiích (obr. 9). Zejména, rostlinný hormon, signální transdukce (2. nejvyšší v S1, 8-nejvyšší v S2, nejvyšší v S3, S4 a S5) a rostlina-patogen interakce (nejvyšší v S1, 3-nejvyšší v S2, 2.-nejvyšší v S3, 4-nejvyšší v S4, a 15-nejvyšší v S5) byly relativně vysoce upregulated ve všech fázích. Phenylpropanoid biosyntézy (není v top 15 v S1, nejvyšší v S2, 3-nejvyšší v S3, 7-nejvyšší v S4, 11-nejvyšší v S5), ubichinon a dalších terpenových-chinon biosyntézy (není v top 15 v S1, 6-nejvyšší v S2, 7-nejvyšší v S3, 2.-nejvyšší v S4, 6-nejvyšší v S5) byly také relativně vysoce upregulated ve všech fázích s výjimkou první. Flavonoidů biosyntéza cesta nebyla vysoce obohacený v raných fázích (není v top 15 v S1, 14-nejvyšší v S2), a vysoce obohaceného v pozdních fázích (3. až 4. nejvyšší během fáze 3-5). (alpha-)Linolové kyseliny metabolismus byl vysoce obohacený ve střední fázi (9.-nejvyšší v S1, 2.-nejvyšší v S2, 5. a 6. nejvyšší v S3, 8-nejvyšší v S4, není v top 15 v S5).

srovnávací analýza obohacení metabolické dráhy KEGG. Top 15 cest, obohacený L. ruthenicum ve srovnání s L. barbarum jsou zobrazeny na levé (červená), a ty, obohacený L. barbarum ve srovnání s L. ruthenicum na pravé (zelené). Vývojové fáze (1-5) jsou uvedeny na obrázku. q-hodnota je p-hodnota upravená FDR.
Mezi cestami downregulated v L. ruthenicum ve srovnání s L. barbarum (Obr. 9) významné změny byly pozorovány mezi raných fázích (1 a 2), když cyanoamino metabolismu kyseliny a karotenoidů biosyntéza jsou většinou vysoce downregulated, a pozdní fáze (4 a 5), když SNARE interakce v vezikulární transport, nikotinát a nikotinamid metabolismus, a porfyrinu a chlorofyl metabolismu byly důsledně relativně vysoce downregulated.
Transcriptome – jednotlivé geny
Mezi nejvíce velmi rozdílně vyjádřené geny, některé z nich byly vývojové fáze-konkrétní (tj. velmi rozdílně regulovány pouze na začátku nebo na konci vývojové fáze), ale některé byly důsledně velmi rozdílně regulovány v průběhu všech pěti studovaných fázích (Doplňující datové sady, S4). Několik imunitu-související geny velmi vysoce upregulated v LR ve srovnání s LB v raných vývojových fázích jsou mezi příklady vývojové fáze-konkrétní vyjádření, vzor: chitinase byl 2. nejvyšší upregulated ° C v S1 (13.43-krát), nejvyšší v S2 (13.89-krát), ale v pozdějších fázích to nebylo STUPŇŮ. Podobně, EIX receptor 1/2 byl také velmi vysoce upregulated v časných stádiích, nejvyšší v S1 (13.70) a 2. nejvyšší v S2 (10.90), ale bylo to také není identifikován jako DEG v pozdějších fázích. Některé souvisejí s růstem geny také vykazoval podobné vyjádření vzor: fosfoglycerát kinázy (PGK) byl mezi hrstkou nejvíce vysoce upregulated genů v prvních třech fázích (13.14, 12.87 a 12.77 respektive), ale v pozdějších stádiích nebyl také identifikován jako DEG. CCR4-NOT transkripce podjednotky komplexu 7/8 (CNOT7/8), také vykazovaly velmi podobné výrazu vzor: vysoce upregulated v prvních třech fázích, a ne DEG ve fázích 4 a 5. Několik flavonoidů a phenylpropanoid biosyntéza-související geny vykazovaly opačný vývojové fáze-konkrétní vyjádření, vzor, s relativně nízkou výraz v raných fázích, a velmi vysoké v pozdějších fázích. Příklady jsou: bifunkční dihydroflavonol 4-reduktázy/flavanone 4-reduktázy (DFR), který byl mírně upregulated v LR v S1 (2.25), není DEG v S2, vysoce upregulated v S3 (7.79), a 3-nejvyšší upregulated v genu S4 (14.25) a S5 (16.03). A paralogue tohoto genu vykazovaly téměř totožný vzor: mírně upregulated v S1 (2.44), není DEG v S2, vysoce upregulated v S3 (7.40), 6-nejvyšší upregulated STUPŇŮ v S4 (13.26) a 5. v S5 (14.59). Podobně, flavonoidů, 3′,5′-hydroxylázy (F3’5′ H) nebyla ° v prvních dvou fázích, vysoce upregulated v S3 (6.69), 5-nejvyšší upregulated v genu S4 (13.42), a 4-nejvyšší v S5 (15.05). Flavonoid O-methyltransferáza (OMT) nebyl STUPŇŮ v S1, ale S2 je již vystavoval střední-vysoká upregulace úrovni (4.32), u S3 to byl již třetí nejvyšší upregulated DEG (13.30), a to byl nejvyšší upregulated v genu S4 (18.73) a S5 (18.10). Leucoanthocyanidin dioxygenase (LDOX; anthocyanin biosyntéza) nebyl STUPŇŮ v S1 a S2, následuje vysoká až velmi vysoká buněk v pozdějších fázích (5.63, 9.44, 11.56, respektive). Dva paralogy chalkonsyntázy (CHS a CHS2; biosyntéza flavonoidů) také nebyly vysoce regulovány v S1 a S2 (CHS2: ne DEG, CHS: -1.14 v S1, ne DEG v S2), ale v S3–S5 oba geny vykazovaly středně vysoká až vysoká upregulace (CHS2: 5.32, 7.84, 6.00; a CHS: 4.67, 7.01, 6.82; respektive). Tyto geny jsme vybrali pro qPCR analýzu a výsledky jsou vysoce shodné s daty RNA-seq (doplňkové výsledky; Doplňkový Dataset S5). Konečně, hodnotami dehydrogenázy, zeatin biosyntéza-related gene, byl také stále upregulovány během posledních tří etap (2.6–5.7).
některé geny však byly konzistentně odlišně exprimovány ve všech pěti studovaných stádiích. Příklady zahrnuty také některé imunity související s geny, jako dva paralogues glutathion S-transferázy, vysoce upregulated v LR ve srovnání s LB ve všech fázích: 9.38 a 8.58 (všechny hodnoty uvedené jako fold-změny v příslušné objednávky) v S1, 6.30 a 6.34 v S2, 2. a 7. nejvyšší upregulated DEGs v S3 (14.08 a 12.70), 2. a 4. nejvyšší v S4 (15.71 a 14.16) a 2. a 6. nejvyšší v S5 (16.48 a 14.40). Choroby rostlin odolnost proti RPM1 protein byl také vysoce upregulated ve všech pěti fází (S1 = 13.15; S2 = 12.08; S3 = 13.11, S4 = 12.81; S5 = 13:94). Mezi důsledně rozdílně vyjádřené genů v průběhu všech vývojových fází byly také některé související s metabolismu aminokyselin, ale jejich vzor byl obrátil v porovnání s předchozími příklady: jsou vystaveny vysoké downregulation v LR ve srovnání s LB. Příklady jsou acetyl-CoA acyltransferase 1 (AAT1; valin, leucin a isoleucin degradace), s temporální profil stále vysoké downregulation, od -7.0 v S1 <−10-krát v posledních třech etapách. Prolin iminopeptidase, spojené s arginin a prolin metabolismus, byl velmi downregulated v LR ve všech fázích: S1 = -9.75, S2 = -10.89 (3. nejvyšší), S3 = -11.05 (4. nejvyšší), S4 = -10.01, a S5 = -11.98 (3. nejvyšší). Konečně, 5-methyltetrahydropteroyltriglutamate–homocystein methyltransferázy (metE) byl důsledně velmi downregulated v LR ve všech fázích: 2.-nejvyšší v S1 (-11.76), nejvyšší v S2 (-11.74), 3-nejvyšší v S3 (-11.43), nejvyšší v S4 (-12.36), a 2. nejvyšší v S5 (-12.83). Dva replikace DNA a transkripce-spojené geny byly také vysoce downregulated v LR ve všech fázích: GTP-vazebný jaderný protein, Ran (RAN; -10.0 až -12.0) a replikační faktor A1 (RFA1) (-8.0 až -12.0). Některé růst a stresu-související geny byly také důsledně vysoce downregulated v LR: heterogenní jaderné ribonucleoprotein A1/A3 (hnRNP; -7 až -11) a tepelného šoku 70 kDa protein 1/8 (HSPA1_8) S1 = -4.95, S2 = -8.88, S3 = -11.48 (2. nejvyšší), S4 = -9.36, S5 = -12.89 (nejvyšší). Je zajímavé, phenylpropanoid biosyntéza-related gene, shikimate hydroxycinnamoyltransferase (HCT), byla také důsledně vysoce downregulated v LR: S1 = -6.82, S2 = -8.14, S3 = -11.71 (nejvyšší), S4 = -11.00 (3. nejvyšší), S5 = -11.91 (4. nejvyšší). Nicméně, klíčový regulátor anthocyanin biosyntéza, transkripční faktor MYB114, byl vysoce upregulated v LR v průběhu všech pět vývojových etap: 6.11, 4.69, 7.47, 9.05, a 8.95 (S1–S5).
metabolické dráhy
provedli jsme také srovnávací mezidruhovou fázovou analýzu metabolických drah (obr. 10). V prvním vývojovém stádiu (S1) jsme identifikovali 39 odlišně regulovaných metabolitů. Mezi top 20 cest tyto metabolity byly spojeny s několika z nich byly spojeny s aminokyselinami, ale celkový počet metabolitů na cestu byly relativně malé (1-2), a P-hodnoty nenaznačují vysokou úroveň významnosti (Obr. 10-panel 1). Zejména metabolismus vitaminu B6 (m) a mikrobiální m v různých prostředích vykazovaly poměrně vysoké hodnoty P, faktor obohacení (EF) 1,0 a 2 identifikované metabolity. V S2, jsme identifikovali 58 diferenciálně regulované metabolitů, spojený s pouze čtyřmi cestami: tryptofan m, phenylpropanoid biosyntézy (b), b phenylpropanoids (to jsou dvě různé cesty v KEGG databáze), a fenylalanin, tyrosin a tryptofan b (všechny EF = 1.0, 2-3 metabolitů, a p < 0.5; Obr. 10-panel 2). V S3, jsme identifikovali 59 diferenciálně regulované metabolitů, spojená s 19 cestami, většinou vše, s EF 1.0, ale relativně non-významné P-hodnoty (>0.5; Obr. 10-panel 3). Cesty s relativně vysokým počtem metabolitů (n = 5) byly: trávení a absorpce bílkovin, B sekundárních metabolitů rostlin, B antibiotik a B aminokyselin. V S4, jsme identifikovali 58 diferenciálně regulované metabolitů, spojené s velkým počtem cest, většinou s EF 1.0, a poměrně vysoký význam hodnot (většinou P > 0.5; Obr. 10-panel 4). Cesty s poměrně vysokým počtem metabolitů (n ≥ 3): phenylpropanoid b, fenylalanin, tyrosin a tryptofan b, glucosinate b, b alkaloidy odvozené od shikimate dráhy, a 2-oxocarboxylic m. V zralého ovoce (S5), jsme identifikovali 39 diferenciálně regulované metabolitů, spojené s velkým počtem drah, ale většinou s nízkou P-hodnoty a pouze 1 metabolitu na dráze (Obr. 10-panel 5). Dráhy s více než 1 metabolitem byly: trávení bílkovin a vstřebávání, phenylpropanoid b, minerální absorpce, centrální uhlík m v rakovinu, b sekundární metabolity, b phenylpropanoids, a aminoacy tRNA b. Analýza hlavních komponent (PCA) všech údajů (druh 2 × 5 etap × 5 biologických replikátů) odhalily vysokou podobnost mezi biologickými opakováními (clustering), a potvrdila významné variability mezi různými zrání ovoce fází pro oba druhy (Obr. 10-panel 6).

srovnávací analýza obohacení metabolické dráhy KEGG. Top 15 cest, obohacený L. ruthenicum ve srovnání s L. barbarum jsou zobrazeny na levé (červená), a ty, obohacený L. barbarum ve srovnání s L. ruthenicum na pravé (zelené). Vývojové fáze (1-5) jsou uvedeny na obrázku. q-hodnota je p-hodnota upravená FDR.
Metabolome – jednotlivé metabolity
seznam (Doplňkové datové sady S6) většina vysoce diferenciálně regulované metabolitů mezi dvěma druhy, vystaveny některé variace mezi pět ovoce fázích vývoje. Je zajímavé, Fruktóza 1-fosfát byl nejvíce vysoce upregulated metabolitu v LR, ve srovnání s LB, během všech pěti etap: log2 Složit změna = 6.3, 7.6, 7.7, 8.1, a 6.5 (fáze 1 až 5 v uvedeném pořadí). Pokud jde o metabolity vysoce upregulované v LB, mezi fázemi bylo více variací: v S1, rozdíly byly poměrně malé, s 9-Decenol jako nejvíce vysoce upregulated metabolit (log2 Složit změna = 2.7; ve srovnání s LR). Analýzy S2 a S3 vyrábí vysoce shodné výsledky, s fenol (3.7 a 3.2 v tomto pořadí) jako nejvíce vysoce upregulated metabolitu. V S4, indoxyl sulfát (4.7) byl vrchol metabolit upregulated v LB. Metabolický posun byl zaznamenán v zralého ovoce (S5), kde seznam metabolitů upregulated v LB byla završena tím, že stearoylcarnitine (7.1), Methoxyoctová kyselina (5.3), S-Methyl-5′-thioadenosine (4.7), lisinopril (4.7), adenosin 3′, 5′ – cyklický fosfát (cAMP) (4.7) atd. Další metabolity vysoce upregulated v LR (kromě Fruktózy 1-fosfát) byla naringin (6.2), – lauroyl-CoA (4.8), L-Phneylalanine (4.6), atd.