keräsimme L. barbarum-ja L. ruthenicum-lajien hedelmiä viidessä kehitysvaiheessa nuorista hedelmistä (≈10 päivää kukinnan jälkeen) täysikasvuisiin (34-45 päivää kukinnan jälkeen) ja tutkimme niiden transkriptomia ja metabolomia.
- RNA-seq de novo assembly and functional annotation of unigenes
- Lycium barbarum (lb): pairwise interstage analyses
- LB Transkriptome
- LB-Metabolomi
- Lycium ruthenicum (LR): pairwise interstage analyses
- LR Transkriptome
- LR-Metabolomi
- Lajienvälinen vertaileva analyysi sukujen eri kehitysvaiheista
- sukujen kokonaismäärä hedelmänkehityksen aikana
- Transkriptome-pathways
- Transkriptomi-yksittäiset geenit
- Metabolomireitit
- Metabolomi-yksittäiset metaboliitit
RNA-seq de novo assembly and functional annotation of unigenes
valmistimme L. barbarumin ja L. ruthenicumin hedelmistä yhteensä 30 cDNA-kirjastoa, joissa on kolme biologista toisintoa (kolme hedelmää kolmesta puusta) kullakin aikapisteellä: 2 lajia × 5 aikapistettä × 3 biologista toisintoa. Näytteille annettiin merkintä LB / LR(1-5)-(1-3), jossa LB on L. barbarum ja LR on L. ruthenicum, 1-5 ovat hedelmien kehitysvaiheita (S1–S5) ja 1-3 ovat yksittäisiä näytteitä (biologisia toisintoja), joten esimerkiksi LB1-1 edustaa L. barbarum, 1st sampled developmental stage (S1), fruit sample No. 1 (out of three). Tuotimme yli 1,72 miljardia paripään lukua näille 30 cDNA-kirjastolle, mikä vastaa keskimäärin 57,2 miljoonaa lukua otosta kohti (täydentävä aineisto S1). Tiukka laadunarviointi ja tietojen suodatus tuotti yhteensä 801 766 laadukasta lukua, joiden keskimääräinen pituus oli 730 ja N50 1107 bp (Taulukko 1). Lopulta transkripteista saatiin yhteensä 326 276 unigeenia, joiden keskimääräinen pituus oli 596 bp ja N50 847 bp (Taulukko 1). 30 näytteen RNA-seq-tietojen korrelaatiokertoimet osoittavat tulosten olevan erittäin yhdenmukaisia biologisten rinnakkaisnäytteiden välillä (Kuva. 2).

Heatmap of correlation coefficients for RNA-seq data for 30 samples of L. barbarum (LB) and L. ruthenicum (LR) fruits at five different development stage. Näytteille annetaan merkintä LB / R1–5_1–3, jossa LB on L. barbarum, LR on L. ruthenicum, 1-5 ovat hedelmien kehitysvaiheita ja 1-3 yksittäistä näytettä. Näytteet ryhmiteltiin hierarkkisen ryhmittelyn mukaan; heatmapin yläpuolella ja vasemmalla puolella olevat dendrogramit osoittavat näytteiden vastaavuuden.
kaikista 326 276 unigeenistä, joita tiedusteltiin julkisista tietokannoista, yhteensä 193 021 (59,15%) täsmäsi geenejä ja/tai proteiineja ainakin yhdessä tietokannassa, ja 12 171 (3,73%) merkittiin kaikkiin tietokantoihin. Eniten unigeenejä (149 863, 45,93%) merkittiin NT-tietokantaan ja vähiten (24 017; 7,36%) KOG-tietokantaan.
Lycium barbarum (lb): pairwise interstage analyses
LB Transkriptome
korkeimmat Degit havaittiin kaikissa 1.vaiheen parivertailuissa ja 2. vs. 5. vaiheen vertailussa (kaikki > 10 000 astetta; Taulukko 2). Pienimmät luvut tunnistettiin 3. vs. 4. ja 4. vs. 5. vaiheen vertailuissa (255-257). Heatmap analyysi DEGs LB osoittaa, että melko erilaiset geenit olivat erittäin upregulated varhaisessa kehitysvaiheessa (1 + 2) ja myöhemmissä vaiheissa (3-5) (Fig. 3 A). Näytteen suhteellisuusanalyysi osoittaa, että näytteet voitaisiin jakaa kahteen kladiin (vaiheet 1 + 2 ja 3 + 4 + 5), jossa jälkimmäinen kladi jaetaan edelleen kahteen kladiin: vaiheisiin 3 + 4 ja vaiheeseen 5. Intraspecific Kegg functional classification analysis of these DEGs identified 15 Trails significantly (p < 0.05) differentially regulated between different developmental stages (Supplementary Dataset S2). Erityisen voimakkaasti toisistaan poikkeavia olivat ”kasvihormonisignaalin transduktio”, ” fenyylipropanoidibiosynteesi (b.)”, ” linolihapon metabolia (m.)”, ” tärkkelys ja sakkaroosi m.”, ja ”zeatin b.” (kuva. 3 A).

Heatmaps and functional Reach analyses of differentially expressed genes (DEGs) in Lycium barbarum (panel A) and L. ruthenicum (panel B) fruits. Heatmaps: t syntyivät hierarkkisella analyysillä DEGs: stä (y-akseli) ja yksittäisistä näytteistä (x-akseli), jossa heatmapin ylä-ja vasemmanpuoleiset dendrografit osoittavat näytteiden relatediteetin. Näytteille annetaan merkintä LB / R_1–5_1-3, jossa lajin kirjainta (LB tai LR) seuraa hedelmän kehitysvaihe (1-5) ja näytteen numero (1-3). Heatmaps: n oikealla puolella on intraspecific KEGG-reittianalyysit DEGs: stä näiden kahden lajin kaikissa viidessä kehitysvaiheessa. Listalla on vain 15 parasta rikastettua reittiä. q-arvo on FDR-mukautettu p-arvo.
aineiston syvällisempää analysointia varten keskityimme vertailemaan merkittävimpiä säädeltyjä reittejä peräkkäisissä kehitysvaiheissa. Ensimmäisessä parivertailussa (S1 vs. S2) ’fenyylipropanoidi b.’ oli eniten toisistaan poikkeava säätelyreitti, jota seurasi ’tärkkelys ja sakkaroosi m.’ (kuva. 4). Hyvin suuri määrä DEGs (>100) todettiin molemmilla poluilla. Sama tulos havaittiin seuraavassa parivertailussa, S2 vs. S3, mutta siitä huolimatta, että Degien (>80) tärkkelys-ja sakkaroosipitoisuudet olivat melko suuria.”Q-arvo on jonkin verran pienempi. S3 vs. S4-vertailussa ”Carbon fixation in photosynthetic, eliöt” oli merkittävimmin säännelty reitti, mutta geenien määrä oli paljon pienempi. Viimeisessä parissa, S4 vs. S5, ” tseatiini b.”, flavonoidi b.”, rasvahappo b. ” ja ”galaktoosi m.” olivat merkittävimmin säännellyt reitit, mutta yhdessäkään reitistä esiintyi enemmän kuin kaksi astetta.

LB transcriptome: pairwise interstage KEGG metaboliatien rikastus. (A) Vaihe 1 vs. Vaihe 2. B) Vaihe 2 vs. Vaihe 3. C) Vaihe 3 vs. Vaihe 4. D) Vaihe 4 vs. vaihe 5. Top 15 (tai kaikki, jos yhteensä <15) merkitsevästi rikastuneet reitit on esitetty. Tangon väri kuvaa Q-arvon suuruutta (FDR-mukautettu p-arvo). Kuvassa näkyy värikartta.
LB-Metabolomi
teimme pareittain kehitysvaihevertailuja tunnistaaksemme metaboliittien rikastumisen kaikkien vaiheparien välillä. LB: n kehitysvaiheiden parivertailuissa erilaisten säänneltyjen metaboliittien määrä vaihteli 66: sta (vaiheet 3 vs. 4) 129: ään (vaiheet 2 vs. 5). Suurin luku peräkkäisissä vaihevertailuissa havaittiin 1.ja 2. vaiheen välillä (117). Näitä osoitettiin suurelle määrälle reittejä; eniten metaboliitteja osoitettiin sekundaarimetaboliittien b-arvolle (myös alhaisin p-arvo), minkä jälkeen tulivat proteiinidigestio ja imeytyminen sekä aminohappojen B-arvo (Kuva. 5; Täydentäviä Lukuja). Peräkkäisessä vaiheparivertailussa (S2 vs. S3), ”ABC transporters” oli merkittävimmin rikastettu reitti, jota seurasi ” purine m.”. S3 vs. S4-parivertailussa merkittävimmin rikastui ”microbial m. in diverse environments”, jota seurasi ” carbapenem m.”. S4 vs. S5-parivertailussa suhteellisen suuri määrä reittejä osoitti samanlaisia tuloksia (kaksi metaboliittia ja samanlaiset p-arvot), mutta huomattava on ”isoflavonoidi b: n” ja ”flavonoidi b: n” esiintyminen niiden joukossa (Kuva. 5; Täydentäviä Lukuja).

LB-metabolomitiedot: pairwise interstage KEGG metaboliatien rikastuminen. (A) Vaihe 1 vs. Vaihe 2. (B) Vaihe 2 vs. Vaihe 3. C) Vaihe 3 vs. Vaihe 4. D) Vaihe 4 vs. vaihe 5. Rikkatekijä on reitissä merkitsevästi säänneltyjen metaboliittien määrän ja reitillä merkittyjen metaboliittien kokonaismäärän suhde (vaihteluväli = 0-1, 0). Pisteen koko kuvaa merkittävästi rikastuneiden metaboliittien määrää vastaavalla reitillä, ja pisteen väri kuvaa P-arvoa (molemmat kuvat kuvassa).
yksittäisistä metaboliiteista S1 vs. S2-vertailussa S1: ssä säädeltyjen metaboliittien luettelon kärjessä olivat trehaloosi, Glaktinoli ja L-omenahappo (kaikki ≈20 log2FC). Öljyhappo, 2-Oksoadipiinihappo ja steariinihappo olivat S2: n eniten sääteleviä metaboliitteja (kaikki ≈15-18 log2FC). S2:n ja S3: n eniten säätelevien metaboliittien listan kärjessä olivat dihydroksiasetoni, LysoPC(18: 1(9Z)) ja adeniini (kaikki ≈16-17, 5 log2FC). S3: n (verrattuna S2: een) eniten säädeltyjen metaboliittien listan kärjessä olivat trehaloosi, Galaktinoli ja L-omenahappo (kaikki ≈19-20 log2FC). S3 vs. S4-vertailussa S3:n eniten säädeltyjen metaboliittien listan kärjessä olivat L-omenahappo, DL-arginiini ja öljyhappo (kaikki ≈18-19 log2FC), kun taas S4: n eniten säädeltyjen metaboliittien listan kärjessä olivat D-mannoosi, N-Asetyllaktosamiini ja LysoPC(18: 1(9Z)) lb: ssä (kaikki ≈16 log2FC). S4 vs. S5 vertailu, S4: n eniten säädeltyjen metaboliittien listan kärjessä olivat trehaloosi, palmitiinihappo, N-Asetyllaktosamiini (kaikki ≈17-18 log2FC) LB: ssä. S5: n eniten säädeltyjen metaboliittien listan kärjessä olivat L-Norleusiini, Antraniilihappo (L1-vitamiini) ja DL-arginiini (kaikki ≈16-18 log2FC) (täydentävä aineisto S3).
Lycium ruthenicum (LR): pairwise interstage analyses
LR Transkriptome
Heatmap analysis of DEGs in LR osoittaa, että lähes täysin erilaiset geenit regulated in the early development stage (1 + 2) and in kypsät hedelmät (stage 5), näennäinen transkriptomic reset tapahtuu jälkeen toisen vaiheen (Fig. 3b). Näytteen suhteellisuusanalyysi osoittaa, että näytteet voitaisiin jakaa kahteen kladiin (vaiheet 1 + 2 ja 3 + 4 + 5), jossa jälkimmäinen kladi jaetaan edelleen kahteen kladiin: vaiheisiin 3 + 4 ja vaiheeseen 5. Intraspecific Kegg functional classification analysis of these DEGs identified 35 Trails significantly (p < 0.05) differentially regulated between different developmental stages (Supplementary Dataset S2). Erityisen voimakkaasti toisistaan poikkeavia olivat ”fotosynteesi”, ” linolihappo m.”, ” isokinoliinialkaloidi b.”, ”flavonoidi b.” ja ”tärkkelys ja sakkaroosi m” (kuva. 3b). Korkeimmat asteiden määrät havaittiin useimmissa vaiheen 1 parivertailuissa (1 vs. 2 oli poikkeus) ja vaiheen 2 vs. 5 vertailussa (kaikki >5000 astetta; Taulukko 2). Ylivoimaisesti pienin luku havaittiin S3 vs. S4-vertailussa (39). S1 vs S2-parivertailussa ’ aminosokeri ja nukleotidisokeri m. ’oli eniten toisistaan poikkeava reitti, jota seurasi’ linolihappo m. ’ (kuva. 6). S2 vs. S3-parivertailussa merkittävimmät toisistaan poikkeavat reitit olivat ” isokinoliinialkaloidi b.”, ” tyrosiini m.”, ”fenyylipropanoidi b” ja ” flavonoidi b.”. Molemmissa parivertailuissa eniten Degejä (>15 ja >35) havaittiin ”tärkkelys ja sakkaroosi m”. S3 vs. S4 vertailu,’ (alfa-)linolihappo m. ’ oli merkittävimmin säännelty reitti, mutta geenien määrä oli paljon pienempi. S4 vs. S5-vertailussa ”fenyylipropanoidi b” (myös eniten Degejä) ja ”linolihappo m.” olivat merkittävimmin säänneltyjä reittejä.
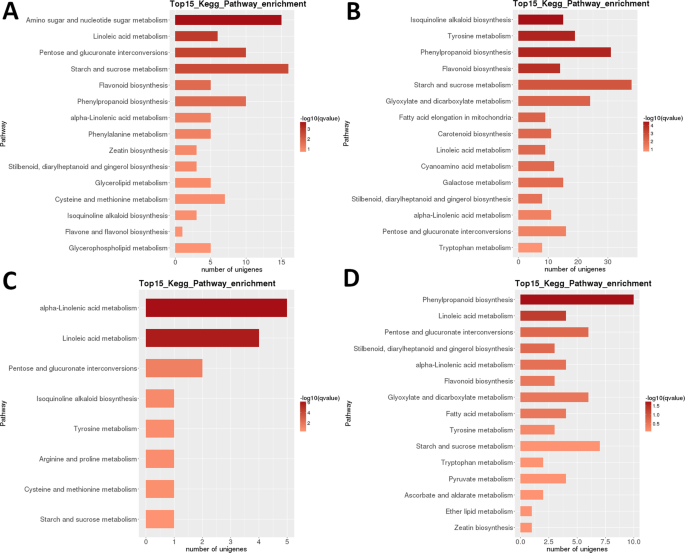
LR transcriptome: pairwise interstage KEGG metabolic pathway enrikastuminen. (A) Vaihe 1 vs. Vaihe 2. (B) Vaihe 2 vs. Vaihe 3. C) Vaihe 3 vs. Vaihe 4. D) Vaihe 4 vs. vaihe 5. Top 15 (tai kaikki, jos yhteensä <15) merkitsevästi rikastuneet reitit on esitetty. Tangon väri kuvaa Q-arvon suuruutta (FDR-mukautettu p-arvo, kuvassa näkyvä värikartta).
LR-Metabolomi
LR-metaboliitin kehitysvaiheiden parivertailuissa eri tavoin säänneltyjen metaboliittien määrä vaihteli 66: sta (vaiheet 3 vs. 4) 133: een (vaiheet 2 vs. 5) (lisäaineisto S3). Peräkkäisissä vaihevertailuissa suurin luku havaittiin 1.ja 2. vaiheen välillä (117). Näitä määrättiin lukuisille poluille; suurin määrä metaboliitteja, jotka on osoitettu sekundaarimetaboliittien b: lle, jota seuraa proteiinien pilkkoutuminen ja imeytyminen, aminohappojen B: lle ja flavonoidi b: lle. S2 vs. S3-vertailussa ”ABC transporters” oli merkittävimmin rikastettu väylä, jota seurasi ”purine m”. S3 vs. S4-parivertailussa merkittävimmin rikastui ”microbial m. in diverse environments”, jota seurasi ” carbapenem m.”. S4 vs. S5-parivertailu, suhteellisen suuri määrä reittejä, joilla oli samanlaisia tuloksia (2 metaboliittia ja samanlaiset p-arvot), mutta huomattava on ”isoflavonoidi b: n” ja ”flavonoidi b: n” esiintyminen niiden joukossa (Kuva. 7; Täydentäviä Lukuja).

LR metabolomitiedot: pairwise interstage KEGG metaboliatien rikastuminen. (A) Vaihe 1 vs. Vaihe 2. (B) Vaihe 2 vs. Vaihe 3. C) Vaihe 3 vs. Vaihe 4. D) Vaihe 4 vs. vaihe 5. Rikkatekijä on reitissä merkitsevästi säänneltyjen metaboliittien määrän ja reitillä merkittyjen metaboliittien kokonaismäärän suhde (vaihteluväli = 0-1, 0). Pisteen koko kuvaa merkittävästi rikastuneiden metaboliittien määrää vastaavalla reitillä, ja pisteen väri kuvaa P-arvoa (molemmat kuvat kuvassa).
yksittäisten metaboliittien (täydentävä tietokokonaisuus S3) osalta S1 vs. S2 vertailu, S1: ssä ylöspäin säädettyjen metaboliittien luettelon kärjessä olivat trehaloosi, Galaktinoli ja L-omenahappo (≈19-21 log2FC), kun taas öljyhappo, 2-Oksoadipiinihappo ja steariinihappo olivat S2: n eniten säädellyt metaboliitit (≈15-18 log2FC). S2 vs. S3-vertailussa dihydroksiasetoni, Indoksyylisulfaatti ja N-Asetyllaktosamiini olivat voimakkaimmin sääteltyjä metaboliitteja S2: ssa (≈17-19, 5 log2FC) ja trehaloosi, Galaktinoli ja L-omenahappo (≈19-21 log2FC) S3: ssa. S3 vs. S4 vertailu, L-omenahappo, DL-arginiini ja öljyhappo olivat upregulated in S3 (≈16-19 log2FC), ja 1,7-Dimetyyliksantiini, D-mannoosi ja N-Asetyllaktosamiini (≈15-17 log2FC) in S4. S4 vs. S5-vertailussa Flaviinimononukleotidi, trehaloosi ja Isoferuliinihappo olivat S4:n (≈18-20 log2FC) suuruisia, kun taas Pg(16:0/18: 1(9Z)), D-proliini ja DL-arginiini olivat voimakkaimmin suurisäätöisiä (kaikki ≈16-18 log2FC) metaboliitteja.
Lajienvälinen vertaileva analyysi sukujen eri kehitysvaiheista
sukujen kokonaismäärä hedelmänkehityksen aikana
Lajienvälinen parivertailu (LR1 vs. LB1, LR2 vs. LB2 jne.) osoittaa, että 928 DEGs oli jaettu kaikki viisi paria(Kuva. 8 A). Korkein määrä DEGs tunnistettiin vaiheessa 3 (3989), ja pienin vaiheessa 4 (2825) (Kuva. 8B); ottaa huomioon, että korkeimmat asteiden määrät, jotka olivat ainutlaatuisia parille, havaittiin vaiheissa 3 (574), 1 ja 5 (molemmat 554) ja alhaisimmat vaiheessa 4 (126) (Kuva. 8 A). Määrä ylös-ja alas-säännelty aste olivat suhteellisen samanlaisia kussakin pairwise vaiheessa vertailut; esim vaiheessa 5, 1668 astetta oli ylös-ja 1670 astetta oli alas-regulated LR verrattuna LB (Kuva. 8B). Neljässä muussa vaiheessa yläsäädeltyjen geenien määrä oli kuitenkin hieman (93-189 astetta) suurempi.

geenit eroavat toisistaan L. barbarumin (lb) ja L. ruthenicumin (LR) hedelmien välillä. (A) yksityiskohtaiset stagewise (1-5) vertailut (LB vs. LR). (B) UPREGULATED (red) and downregulated (green) DEGs in LR verrattuna LB viidessä tutkitussa kehitysvaiheessa.
Transkriptome-pathways
Heatmap-analyysi DEGs: stä osoittaa, että kahden lajin hedelmillä on hyvin erilaiset geeniekspressioprofiilit kaikissa kehitysvaiheissa, mutta biologisilla toisinnoilla oli hyvin samanlaiset profiilit, mikä viittaa rajalliseen yksilökohtaiseen vaihteluun kussakin kehitysvaiheessa (täydentävät luvut: Fig. S9). Kegg-reitin rikastumisen vertaileva analyysi osoittaa, että vain jotkut reitit olivat johdonmukaisesti erittäin rikastuneita (geenien säätelyn kannalta) LR: ssä verrattuna LB: hen kaikissa viidessä kehitysvaiheessa (Kuva. 9). Erityisesti kasvien hormonisignaalin transduktio (2.korkein S1: ssä, 8. korkein S2: ssa, korkein S3: ssa, S4: ssä ja S5: ssä) ja kasvien ja patogeenien vuorovaikutus (korkein S1: ssä, 3. korkein S2: ssa, 2. korkein S3: ssa, 4. korkein S4: ssä ja 15. korkein S5: ssä) olivat suhteellisen korkeasääntelyä kaikissa vaiheissa. Fenyylipropanoidien biosynteesi (ei top 15: ssä S1: ssä, korkein S2: ssa, 3.korkein S3: ssa, 7. korkein S4: ssä, 11. korkein S5: ssä), ubikinoni ja muut terpenoidikinonin biosynteesi (ei top 15: ssä S1: ssä, 6. korkein S2: ssa, 7. korkein S3: ssa, 2. korkein S4: ssä, 6. korkein S5: ssä) olivat myös suhteellisen korkeasääntelyisiä kaikissa vaiheissa ensimmäistä lukuun ottamatta. Flavonoidien biosynteesireitti ei ollut pitkälle rikastunut alkuvaiheessa (ei Top 15: ssä S1: ssä, 14.korkein S2: ssa) ja pitkälle rikastunut loppuvaiheessa (3. -4. korkein vaiheissa 3-5). (alfa-)linolihapon metabolia rikastui voimakkaasti keskivaiheilla (9.korkein S1: ssä, 2. korkein S2: ssa, 5. ja 6. korkein S3: ssa, 8. korkein S4: ssä, ei top 15: ssä S5: ssä).

vertaileva analyysi keggin metaboliareitin rikastumisesta. L. ruthenicumissa rikastetut 15 parasta reittiä L. barbarumiin verrattuna näkyvät vasemmalla (punainen) ja L. barbarumissa rikastetut L. ruthenicumiin verrattuna oikealla (vihreä). Kehitysvaiheet (1-5) on esitetty kuvassa. q-arvo on FDR-mukautettu p-arvo.
muun muassa L. ruthenicumissa L. barbarumiin verrattavia reittejä (Kuva. 9) merkittäviä muutoksia havaittiin alkuvaiheiden (1 ja 2) välillä, kun syanoaminohapon aineenvaihdunta ja karotenoidibiosynteesi olivat eniten downregulated, ja myöhäisissä vaiheissa (4 ja 5), kun SNARE yhteisvaikutukset vesicular liikenne, nikotinaatti ja nikotiiniamidi aineenvaihduntaa, ja porfyriini ja klorofylli aineenvaihduntaa olivat johdonmukaisesti suhteellisen hyvin downregulated.
Transkriptomi-yksittäiset geenit
kaikkein eniten toisistaan eroavista geeneistä osa oli kehitysastepesifisiä (eli hyvin erilaisia vain varhaisessa tai myöhäisessä kehitysvaiheessa), mutta osa oli johdonmukaisesti hyvin erilaisia kaikissa viidessä tutkitussa vaiheessa (täydentävä aineisto S4). Useat immuniteettiin liittyvät geenit, jotka ovat LR: ssä hyvin korkeasäätöisiä verrattuna lb: hen varhaisessa kehitysvaiheessa, ovat esimerkkejä kehitysvaihespesifisestä ekspressiokuviosta: kitinase oli 2.korkein aste S1: ssä (13,43-kertainen), korkein S2: ssa (13,89-kertainen), mutta myöhemmissä vaiheissa se ei ollut astetta. Samoin EIX-reseptori 1/2 oli myös hyvin korkeasäänteinen alkuvaiheessa, korkein S1: ssä (13,70) ja 2.korkein S2: ssa (10,90), mutta sitä ei myöskään tunnistettu DEGIKSI myöhemmissä vaiheissa. Joillakin kasvuun liittyvillä geeneillä oli myös samanlainen ilmentymämalli: fosfoglyseraattikinaasi (PGK) oli kourallisen eniten ylisääntyneitä geenejä kolmessa ensimmäisessä vaiheessa (13.14, 12.87 ja 12.77), mutta sitä ei myöskään myöhemmissä vaiheissa tunnistettu DEGIKSI. CCR4-NOT transkription complex alayksikkö 7/8 (CNOT7 / 8) oli myös hyvin samanlainen lauseke malli: erittäin ylös säädelty kolmessa ensimmäisessä vaiheessa, eikä astetta vaiheissa 4 ja 5. Useilla flavonoidi-ja fenyylipropanoidibiosynteesiin liittyvillä geeneillä esiintyi käänteistä kehitysastespesifistä ilmentymismallia, jonka ilmentyminen oli alkuvaiheessa suhteellisen vähäistä ja myöhemmissä vaiheissa erittäin suurta. Esimerkkejä ovat: bifunktionaalinen dihydroflavonoli-4-reduktaasi/flavanoni-4-reduktaasi (DFR), joka oli LR: ssä hieman ylöspäin S1: ssä (2.25), ei astetta S2, erittäin ylös regulated S3 (7.79), ja 3rd korkein ylös regulated geeni S4 (14.25) ja S5 (16.03). Tämän geenin paralogi oli lähes identtinen: hieman yläsääntelyä S1: ssä (2.44), Ei astetta S2: ssa, erittäin yläsääntelyä S3: ssa (7.40), 6.korkeinta YLÄSÄÄNTELYÄ S4: ssä (13.26) ja 5. tasoa S5: ssä (14.59). Vastaavasti flavonoidi 3′, 5′-hydroksylaasi (F3 ’ 5 ’ h) ei ollut DEGI kahdessa ensimmäisessä vaiheessa, hyvin ylös reguloitu S3: ssa (6.69), 5.korkein ylös reguloitu geeni S4: ssä (13.42) ja 4. korkein S5: ssä (15.05). Flavonoidi O-metyylitransferaasi (OMT) ei ollut aste S1: ssä, mutta S2: ssa sillä oli jo keskikorkea yläsääntelytaso (4.32), S3: ssa se oli jo kolmanneksi korkein yläsääntelytaso (13.30), ja se oli korkein yläsääntelytaso S4: ssä (18.73) ja S5: ssä (18.10). Leukoantosyanidiinidioksigenaasi (LDOX; antosyaanin biosynteesi) ei ollut aste S1: ssä ja S2: ssa, jota seurasi korkea tai erittäin korkea upregulaatio myöhemmissä vaiheissa (5, 63, 9, 44, 11, 56, vastaavasti). Kaksi kalkonisyntaasi paralogia (CHS ja CHS2; flavonoidien biosynteesi) eivät myöskään olleet kovin säänneltyjä S1: ssä ja S2: ssa (CHS2: Ei astetta, CHS: – 1.14 S1: ssä, ei astetta S2: ssa), mutta S3–S5: ssä molemmilla geeneillä oli keskikorkea tai korkea yläsäätely (CHS2: 5.32, 7.84, 6.00; ja CHS: 4.67, 7.01, 6.82; vastaavasti). Valitsimme nämä geenit qPCR-analyysiin, ja tulokset ovat hyvin yhdenmukaisia RNA-seq-datan kanssa (täydentäviä tuloksia; täydentäviä tietoja S5). Lopulta myös sytokiniinidehydrogenaasi, joka on tseatiinin biosynteesiin liittyvä geeni, oli viimeisen kolmen vaiheen aikana (2,6-5,7) enenevässä määrin ylisääntynyt.
jotkin geenit kuitenkin ilmentyivät johdonmukaisesti eri tavoin kaikissa viidessä tutkitussa vaiheessa. Esimerkkejä olivat myös eräät immuniteettiin liittyvät geenit, kuten kaksi glutationi-s-transferaasin paralogia, joiden LR-arvo oli kaikissa vaiheissa erittäin korkea verrattuna LB: hen: 9.38 ja 8.58 (kaikki arvot esitetty kertamuutoksina vastaavassa järjestyksessä) S1: ssä, 6.30 ja 6.34 S2: ssa, 2.ja 7. korkein S3: ssa (14.08 ja 12.70), 2. ja 4. korkein S4: ssä (15.71 ja 14,16) ja S5: n 2. ja 6. paras (16,48 ja 14,40). Myös kasvitautien resistenssiproteiini RPM1: n säätely oli erittäin korkealla kaikissa viidessä vaiheessa (S1 = 13, 15; S2 = 12, 08; S3 = 13, 11; S4 = 12, 81; S5 = 13:94). Johdonmukaisesti eri tavoin ilmaistujen geenien joukossa kaikissa kehitysvaiheissa oli myös joitakin aminohappometaboliaan liittyviä geenejä, mutta niiden rakenne oli päinvastainen verrattuna aiempiin esimerkkeihin: niillä oli LR: n suuri alasäätely verrattuna LB: hen. Esimerkkejä ovat asetyyli-CoA-asyylitransferaasi 1 (AAT1; valiinin, leusiinin ja isoleusiinin hajoaminen), jonka ajallinen profiili on yhä suurempi, alkaen -7, 0: sta S1: ssä <−10-kertaiseksi kolmessa viimeisessä vaiheessa. Proliini-iminopeptidaasi, joka liittyi arginiinin ja proliinin metaboliaan, laski LR: n säätelyä voimakkaasti kaikissa vaiheissa: S1 = -9, 75, S2 = -10, 89 (3.korkein), S3 = -11, 05 (4. korkein), S4 = -10, 01 ja S5 = -11, 98 (3. korkein). Lopuksi 5–metyylitetrahydropteroyylitriglutamaatti-homokysteiinimetyylitransferaasi (metE) oli jatkuvasti erittäin voimakkaasti alenevassa asemassa LR: ssä kaikissa vaiheissa: 2.korkein S1: ssä (-11.76), korkein S2: ssa (-11.74), 3. korkein S3: ssa (-11.43), korkein S4: ssä (-12.36) ja 2. korkein S5: ssä (-12.83). Myös kaksi DNA: n replikaatioon ja transkriptioon liittyvää geeniä oli LR: ssä kaikissa vaiheissa voimakkaasti alenevassa järjestyksessä: GTP: tä sitova ydinproteiini Ran (RAN;-10, 0- – 12, 0) ja replikaatiotekijä A1 (RFA1) (- 8, 0 – – 12, 0). Jotkin kasvuun ja stressiin liittyvät geenit olivat myös LR: ssä johdonmukaisesti hyvin alenevassa järjestyksessä: heterogeeninen ydinribonukleoproteiini A1/A3 (hnRNP; -7–11) ja lämpöshokki 70 kDa-proteiini 1/8 (HSPA1_8) S1 = -4.95, S2 = -8.88, S3 = -11.48 (2.korkein), S4 = -9.36, S5 = -12.89 (korkein). Mielenkiintoista on, että fenyylipropanoidibiosynteesiin liittyvä geeni, shikimaattihydroksikinnamoyylitransferaasi (HCT), oli myös johdonmukaisesti hyvin alenevassa järjestyksessä LR: ssä: S1 = -6, 82, S2 = -8, 14, S3 = -11, 71 (korkein), S4 = -11, 00 (3.korkein), S5 = -11, 91 (4. korkein). Antosyaanin biosynteesin keskeinen säätelijä, transkriptiotekijä MYB114, oli kuitenkin LR: ssä erittäin korkeasäätöinen kaikissa viidessä kehitysvaiheessa: 6, 11, 4, 69, 7, 47, 9, 05 ja 8, 95 (S1–S5).
Metabolomireitit
teimme myös vertailevan spesifisen vaiheen analyysin metaboliareiteistä (Kuva. 10). Ensimmäisessä kehitysvaiheessa (S1) tunnistimme 39 eri tavalla säänneltyä metaboliittia. Näiden metaboliittien 20 tärkeimmän reitin joukossa useat niistä liittyivät aminohappoihin, mutta metaboliittien kokonaismäärä reittiä kohti oli suhteellisen pieni (1-2), eivätkä P-arvot viitanneet suureen merkitsevyyteen (Fig. 10-paneeli 1). Erityisesti B6-vitamiinin metaboliassa (m) ja mikrobien M: ssä erilaisissa ympäristöissä molemmilla oli verrattain korkeat P-arvot, rikastuskerroin (EF) 1, 0 ja 2 tunnistettua metaboliittia. S2: ssa tunnistimme 58 eri tavalla säänneltyä metaboliittia, jotka liittyvät vain neljään reitiin: tryptofaani m, fenyylipropanoidibiosynteesi (b), fenyylipropanoidien b (Nämä ovat kaksi eri reittiä KEGG-tietokannassa) ja fenyylialaniini, tyrosiini ja tryptofaani b (kaikki EF = 1, 0, 2-3 metaboliittia ja P < 0, 5; Fig. 10-paneeli 2). S3: ssa tunnistimme 59 eri tavalla säänneltyä metaboliittia, jotka liittyvät 19 reitille, useimmat EF 1, 0: lla, mutta suhteellisen ei-merkitsevillä P-arvoilla (>0, 5; Fig. 10-paneeli 3). Metaboliittien suhteellisen suuren määrän (N = 5) reitit olivat seuraavat: proteiinien pilkkoutuminen ja imeytyminen, kasvien sekundaarimetaboliittien b, antibioottien B ja aminohappojen B. S4: ssä tunnistimme 58 eri tavoin säänneltyä metaboliittia, jotka liittyvät suureen määrään reittejä, enimmäkseen EF 1.0: aan, ja verrattain korkeat merkitsevyysarvot (enimmäkseen P > 0.5; Fig. 10-paneeli 4). Reitit, joilla oli suhteellisen suuri määrä metaboliitteja (n ≥ 3), olivat: fenyylipropanoidi b, fenyylialaniini, tyrosiini ja tryptofaani b, glukosinaatti b, shikimaattireitin kautta johdettujen alkaloidien B ja 2-oksokarboksyylinen m. kypsässä hedelmässä (S5) tunnistimme 39 eri tavalla säänneltyä metaboliittia, jotka liittyvät suureen määrään reittejä, mutta enimmäkseen alhaisiin P-arvoihin ja vain 1 metaboliitin per reitti (Fig. 10-paneeli 5). Reitit, joissa oli enemmän kuin 1 metaboliitti, olivat: proteiinin pilkkoutuminen ja imeytyminen, fenyylipropanoidi b, mineraalien imeytyminen, keskeinen hiili m syövässä, sekundaarimetaboliittien b, fenyylipropanoidien b ja aminoaktiivisuus tRNA b.kaikkien tietojen (2 lajia × 5 vaihetta × 5 biologista toisintoa) pääkomponenttianalyysi (PCA) paljasti suuren samankaltaisuuden biologisten toisintojen välillä (ryhmittely) ja vahvisti merkittävää vaihtelua eri hedelmien kypsymisvaiheiden välillä molemmilla lajeilla (Kuva. 10-Paneeli 6).

vertaileva analyysi keggin metaboliareitin rikastumisesta. L. ruthenicumissa rikastetut 15 parasta reittiä L. barbarumiin verrattuna näkyvät vasemmalla (punainen) ja L. barbarumissa rikastetut L. ruthenicumiin verrattuna oikealla (vihreä). Kehitysvaiheet (1-5) on esitetty kuvassa. q-arvo on FDR-mukautettu p-arvo.
Metabolomi-yksittäiset metaboliitit
luettelo (täydentävä tietokokonaisuus S6), jossa on eniten toisistaan poikkeavia säänneltyjä metaboliitteja näiden kahden lajin välillä ja jossa esiintyy jonkin verran vaihtelua hedelmien viiden kehitysvaiheen välillä. Mielenkiintoista on, että fruktoosi-1-fosfaatti oli LR: n eniten säätelevä metaboliitti verrattuna LB: hen kaikissa viidessä vaiheessa: log2-kertainen muutos = 6, 3, 7, 7, 8, 1 ja 6, 5 (vaiheet 1-5). LB: ssä voimakkaasti lisääntyneiden metaboliittien osalta eri vaiheiden välillä oli enemmän vaihtelua: S1: ssä erot olivat melko pieniä, ja 9-Dekenoli oli eniten säätelevä metaboliitti (log2-kertainen muutos = 2, 7; verrattuna LR: ään). S2: n ja S3: n analyysit tuottivat hyvin yhdenmukaisia tuloksia, ja fenoli (3, 7 ja 3, 2) oli eniten säätelevä metaboliitti. S4-ryhmässä INDOKSYYLISULFAATTI (4, 7) oli LB: n päämetaboliitti. Metabolinen muutos havaittiin kypsissä hedelmissä (S5), joissa LB: n säätelemien metaboliittien luettelon kärjessä olivat stearoyylikarnitiini (7.1), Metoksietikkahappo (5.3), S-metyyli-5′-tioadenosiini (4.7), lisinopriili (4.7), adenosiini-3′, 5′ – syklinen fosfaatti (cAMP) (4.7) jne. Muita LR: ssä voimakkaasti esiintyviä metaboliitteja (fruktoosi-1-fosfaattia lukuun ottamatta) olivat naringiini (6.2), lauroyyli-CoA (4.8), L-Fneyylialaniini (4.6) jne.