am colectat fructe de L. barbarum și L. ruthenicum în cinci stadii de dezvoltare, de la fructe tinere (10 zile după înflorire) la fructe mature (34-45 zile după înflorire) și am studiat transcriptomul.
- ARN-seq de novo asamblare și adnotare funcțională a unigenelor
- Lycium barbarum (LB): analize interetajate în perechi
- transcriptom LB
- Metabolomul LB
- Lycium ruthenicum (LR): analizele interetajate în perechi
- transcriptom LR
- Metabolomul LR
- analiza comparativă interspecifică a DEGs în diferite stadii de dezvoltare
- Total DEGs în timpul dezvoltării fructelor
- Transcriptome-pathways
- Transcriptome – Gene individuale
- căi Metabolomice
- metabolom-metaboliți individuali
ARN-seq de novo asamblare și adnotare funcțională a unigenelor
am pregătit un total de 30 de biblioteci ADNc din fructe de L. barbarum și L. ruthenicum, cu trei replici biologice (trei fructe din trei arbori) la fiecare punct de timp: 2 specii 5 puncte de timp 3 exemplare biologice. Probele au fost etichetate LB/LR(1-5)-(1-3), unde LB este L. barbarum și LR este L. ruthenicum, 1-5 sunt stadii de dezvoltare ale fructelor (S1–S5) și 1-3 sunt probe individuale (replici biologice); deci, de exemplu, LB1-1 reprezintă L. barbarum, Etapa 1 de dezvoltare eșantionată (S1), proba de fructe Nr.1 (din trei). Am generat peste 1,72 miliarde de citiri pereche pentru aceste 30 de biblioteci ADNc, corespunzând unei medii de 57,2 milioane de citiri pe eșantion (set de date suplimentar S1). Evaluarea strictă a calității și filtrarea datelor au dat un total de 801.766 de citiri de înaltă calitate, cu lungimea medie de 730 și N50 de 1107 bp (Tabelul 1). În cele din urmă, un total de 326.276 unigeni cu lungimea medie de 596 bp și N50 de 847 bp au fost obținute din transcrieri (Tabelul 1). Coeficienții de corelație pentru ARN-seq datele pentru cele 30 de probe indică o consistență foarte bună a rezultatelor în rândul replicilor biologice (Fig. 2).

harta de căldură a coeficienților de corelație pentru datele ARN-seq pentru 30 de probe de fructe L. barbarum (LB) și L. ruthenicum (LR) în cinci stadii diferite de dezvoltare. Probele sunt etichetate LB/R1–5_1-3, unde LB este L. barbarum, LR este L. ruthenicum, 1-5 sunt etape de dezvoltare a fructelor și 1-3 probe individuale. Probele au fost grupate prin grupare ierarhică; dendrogramele de mai sus și din stânga hărții termice indică corelarea probelor.
dintre toate cele 326.276 de unigene interogate împotriva bazelor de date publice, un total de 193.021 (59,15%) gene și/sau proteine potrivite în cel puțin o bază de date și 12.171 (3,73%) au fost adnotate în toate bazele de date. Cel mai mare număr de unigeni (149.863, 45,93%) a fost adnotat în baza de date NT, iar cel mai mic număr (24.017; 7,36%) în baza de date KOG.
Lycium barbarum (LB): analize interetajate în perechi
transcriptom LB
cel mai mare număr de grade au fost identificate în toate comparațiile în perechi ale etapei 1 și în comparația etapei 2 vs.5 (toate > 10.000 grade; Tabelul 2). Cele mai mici numere au fost identificate în comparațiile etapelor a 3-a VS.A 4-a și a 4-a VS. A 5-a (255-257). Analiza hărții termice a DEGs în LB arată că seturi destul de diferite de gene au fost puternic reglate în stadiile incipiente de dezvoltare (1 + 2) și în etapele ulterioare (3 până la 5) (Fig. 3A). Analiza relaționării eșantioanelor indică faptul că eșantioanele ar putea fi împărțite în două clade (etapele 1 + 2 și 3 + 4 + 5), cu ultima cladă subdivizată în continuare în două clade: etapele 3 + 4 și etapa 5. Analiza clasificării funcționale Kegg intraspecifice a acestor DEGs a identificat 15 căi semnificativ (P < 0,05) reglementate diferențiat între diferite etape de dezvoltare (set de date suplimentar S2). Deosebit de puternic diferențiate au fost ‘transducția semnalului hormonului vegetal’, ‘biosinteza fenilpropanoid (b.)’, ‘metabolismul acidului linoleic (m.)’, ‘ amidonul și zaharoza m.’, și ‘zeatin b.’ (Fig. 3A).

hărți de căldură și analize ale căilor funcționale ale genelor exprimate diferențiat (DEGs) în fructele Lycium barbarum (Panoul a) și L. ruthenicum (panoul B). Hărțile de căldură au fost generate de o analiză ierarhică a DEGs (axa y) și a probelor individuale (axa x), unde dendrogramele de deasupra și din stânga hărții de căldură indică corelarea probelor. Probele sunt etichetate LB / R_1-5_1-3, unde acronimul speciei (LB sau LR) este urmat de stadiul de dezvoltare al fructului (1-5) și de numărul eșantionului (1-3). Analizele intraspecifice ale căii KEGG ale DEGs în toate cele cinci etape de dezvoltare la cele două specii sunt prezentate în partea dreaptă a hărților termice. Sunt listate doar primele 15 căi îmbogățite. valoarea q este o valoare p ajustată FDR.
pentru o analiză mai aprofundată a datelor, ne-am concentrat pe compararea celor mai semnificativ reglementate căi în etapele de dezvoltare succesive. În prima comparație în perechi (S1 vs S2),’ fenilpropanoidul B. ‘a fost calea cea mai diferențiată reglementată, urmată de’ amidon și zaharoză m. ‘ (Fig. 4). Un număr foarte mare de DEGs (>100) au fost identificate în ambele căi. Un rezultat similar a fost observat în următoarea comparație în perechi, S2 vs. S3, dar în ciuda numărului destul de mare de DEGs (>80) ‘amidon și zaharoză m.’au prezentat o valoare q ceva mai mică. În comparația S3 vs. S4, ‘fixarea carbonului în organismele fotosintetice’ a fost calea cea mai semnificativ reglementată, dar numărul de gene a fost mult mai mic. În ultima pereche, S4 vs. S5, ‘zeatin b.’, flavonoid B.’, acid gras b. ‘și’ galactoză m. ‘ au fost căile cele mai semnificativ reglementate, dar niciuna dintre căi nu a prezentat mai mult de două grade.

transcriptom LB: îmbogățirea căii metabolice Kegg în perechi între etape. (A) Etapa 1 vs.etapa 2. (B) Etapa 2 vs. Etapa 3. (C) Etapa 3 vs.etapa 4. (D) Etapa 4 vs.etapa 5. Top 15 (sau toate dacă sunt totale <15) sunt afișate căi semnificativ îmbogățite. Culoarea barei reprezintă magnitudinea valorii q (o valoare p ajustată FDR). Diagrama de culori este prezentată în figură.
Metabolomul LB
am efectuat comparații în etape de dezvoltare în perechi pentru a identifica îmbogățirea metaboliților între toate perechile de etape. Numărul metaboliților reglementați diferențiat în comparațiile perechi ale stadiilor de dezvoltare în LB a variat de la 66 (etapele 3 vs 4) la 129 (etapele 2 vs 5). Cel mai mare număr în comparații succesive de etape a fost observat între etapa 1 și etapa a 2-a (117). Aceștia au fost repartizați pe un număr mare de căi; cu cel mai mare număr de metaboliți repartizați pe ‘b. metaboliților secundari’ (de asemenea, cea mai mică valoare p), urmată de ‘digestia și absorbția proteinelor’ și ‘b. aminoacizilor’ (Fig. 5; Cifre Suplimentare). În comparația succesivă a perechilor de etape (S2 vs. S3),’ ABC transportors ‘a fost calea cea mai semnificativ îmbogățită, urmată de’purine m.’. În comparația perechilor S3 vs. S4, ‘microbian m. în medii diverse’ a fost calea cea mai semnificativ îmbogățită, urmată de ‘carbapenem m.’. În comparația perechilor S4 vs. S5, un număr relativ mare de căi au prezentat rezultate similare (doi metaboliți și valori p similare), dar notabilă este apariția ‘izoflavonoidului b.’ și ‘flavonoidului B.’ printre acestea (Fig. 5; Cifre Suplimentare).

date metabolomice LB: îmbogățirea căii metabolice Kegg interetajate în perechi. (A) Etapa 1 vs.etapa 2. (B) Etapa 2 vs.Etapa 3. (C) Etapa 3 vs.etapa 4. (D) Etapa 4 vs.etapa 5. Factorul bogat este raportul dintre numărul de metaboliți reglementați semnificativ pe cale și numărul total de metaboliți adnotați pe acea cale (interval = 0 până la 1,0). Mărimea punctului reprezintă numărul de metaboliți semnificativ îmbogățiți în calea corespunzătoare, iar culoarea punctului reprezintă valoarea P (ambele legende prezentate în figură).
în ceea ce privește metaboliții individuali, în comparația S1 vs.S2, lista metaboliților regulați în sus în S1 a fost completată de trehaloză, Glactinol și acid L-Malic (toți cei din grupul S20 log2fc). Acidul Oleic, acidul 2-Oxoadipic și acidul Stearic au fost metaboliții cu cea mai mare reglare în S2 (toți 15-18 log2FC). Lista metaboliților cu cea mai mare reglementare în S2 comparativ cu S3 a fost completată de dihidroxiacetonă, Lizopc(18:1(9Z)) și adenină (toate de la 16 la 17,5 log2FC). Lista metaboliților cu cea mai mare reglementare în S3 (în comparație cu S2) a fost completată de trehaloză, Galactinol și acid L-Malic (toate de la 19 la 20 log2FC). În comparația S3 vs. S4, lista celor mai mulți metaboliți cu reglaj ridicat din S3 a fost completată de acid L-Malic, DL-arginină și acid Oleic (toți 18-19 log2FC), în timp ce lista celor mai mulți metaboliți cu reglaj ridicat din S4 a fost completată de D-manoză, n-Acetilactozamină și Lizopc(18:1(9Z)) în LB (toți 16 log2fc). În S4 vs. Comparația S5, lista celor mai mulți metaboliți foarte reglați în S4 a fost depășită de trehaloză, acid Palmitic, n-Acetilactozamină (toate de la 17 la 18 log2FC) în LB. Lista metaboliților cu cea mai mare reglementare în S5 a fost completată de l-Norleucină, acid antranilic (vitamina L1) și DL-arginină (toate de la 16 la 18 log2FC) (set de date suplimentare S3).
Lycium ruthenicum (LR): analizele interetajate în perechi
transcriptom LR
analiza hărții termice a DEGs în LR arată că seturi aproape complet diferite de gene au fost puternic reglate în stadiile incipiente de dezvoltare (1 + 2) și în fructele coapte (etapa 5), cu o resetare transcriptomică aparentă care apare după a doua etapă (Fig. 3B). Analiza relaționării eșantioanelor indică faptul că eșantioanele ar putea fi împărțite în două clade (etapele 1 + 2 și 3 + 4 + 5), cu ultima cladă subdivizată în continuare în două clade: etapele 3 + 4 și etapa 5. Analiza clasificării funcționale Kegg intraspecifice a acestor DEGs a identificat 35 de căi semnificativ (P < 0,05) reglementate diferențiat între diferite etape de dezvoltare (set de date suplimentar S2). Deosebit de puternic diferențiate au fost ‘fotosinteza’, ‘acidul linoleic m.’, ‘alcaloidul izochinolinic b.’, ‘flavonoidul B.’ și ‘amidonul și zaharoza m’ (Fig. 3B). Cel mai mare număr de DEGs a fost identificat în majoritatea comparațiilor în perechi ale etapei 1 (1 vs.2 a fost o excepție), iar în etapa 2 vs. 5 comparație (Toate >5.000 DEGs; Tabelul 2). De departe, cel mai mic număr a fost identificat în comparația S3 vs.S4 (39). În comparația S1 vs S2 în perechi, ‘zahărul amino și zahărul nucleotidic m.’ a fost calea cea mai diferențiată reglementată, urmată de ‘acidul linoleic m.’ (Fig. 6). În comparația perechilor S2 vs. S3, cele mai semnificativ diferențiate căi reglementate au fost ‘alcaloidul izochinolinic b.’, ‘tirozina m.’, ‘fenilpropanoidul B’ și ‘flavonoidul b.’. În ambele comparații în perechi, cel mai mare număr de grade Celsius (>15 și, respectiv, >35) a fost identificat în ‘amidon și zaharoză m.’. În S3 vs. S4 comparație, ‘(alfa-)linoleic acid m. ‘ a fost calea cea mai semnificativ reglementată, dar numărul de gene au fost mult mai mici. În comparația S4 vs. S5, ‘fenilpropanoidul b ‘(de asemenea, cel mai mare număr de DEGs) și’ acidul linoleic m. ‘ au fost căile cele mai semnificativ reglementate.
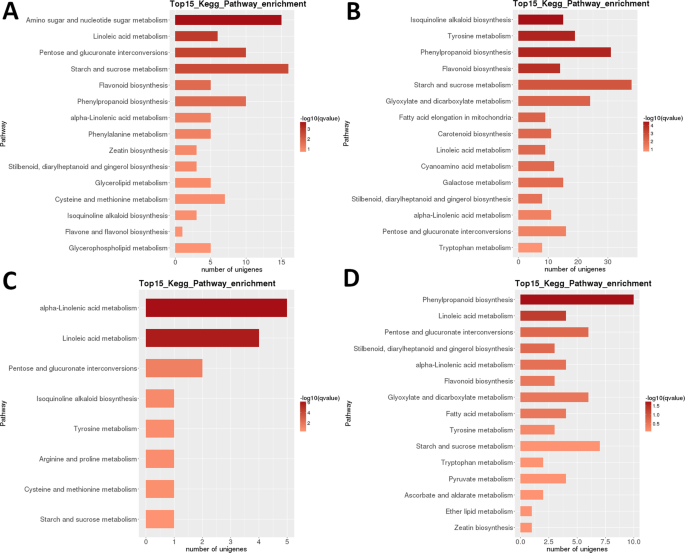
transcriptom LR: îmbogățirea căii metabolice Kegg în perechi între etape. (A) Etapa 1 vs.etapa 2. (B) Etapa 2 vs.Etapa 3. (C) Etapa 3 vs.etapa 4. (D) Etapa 4 vs.etapa 5. Top 15 (sau toate dacă sunt totale <15) sunt afișate căi semnificativ îmbogățite. Culoarea barei reprezintă magnitudinea valorii q (o valoare p ajustată FDR, diagramă de culori prezentată în figură).
Metabolomul LR
Numărul metaboliților reglementați diferențiat în comparațiile perechi ale stadiilor de dezvoltare în LR a variat de la 66 (stadiile 3 vs 4) la 133 (etapele 2 vs 5) (setul de date suplimentar S3). În comparații succesive de etape, cel mai mare număr a fost observat între etapele 1 și 2 (117). Acestea au fost atribuite unui număr mare de căi; cu cel mai mare număr de metaboliți atribuiți metaboliților secundari, urmați de digestia și absorbția proteinelor, aminoacizii și flavonoidul B. În comparația S2 vs. S3, ‘transportatorii ABC ‘a fost calea cea mai semnificativ îmbogățită, urmată de’Purina m’. În comparația perechilor S3 vs. S4, ‘microbian m. în medii diverse’ a fost calea cea mai semnificativ îmbogățită, urmată de ‘carbapenem m.’. În S4 vs. Comparația perechilor S5, un număr relativ mare de căi au prezentat rezultate similare (2 metaboliți și valori p similare), dar notabilă este apariția ‘izoflavonoidului b.’ și ‘flavonoidului B.’ printre acestea (Fig. 7; Cifre Suplimentare).

date metabolomice LR: îmbogățirea căii metabolice Kegg interetajate în perechi. (A) Etapa 1 vs.etapa 2. (B) Etapa 2 vs.Etapa 3. (C) Etapa 3 vs.etapa 4. (D) Etapa 4 vs.etapa 5. Factorul bogat este raportul dintre numărul de metaboliți reglementați semnificativ pe cale și numărul total de metaboliți adnotați pe acea cale (interval = 0 până la 1,0). Mărimea punctului reprezintă numărul de metaboliți semnificativ îmbogățiți în calea corespunzătoare, iar culoarea punctului reprezintă valoarea P (ambele legende prezentate în figură).
în ceea ce privește metaboliții individuali (setul de date suplimentare S3), în S1 vs. Comparația S2, lista metaboliților regulați în sus în S1 a fost completată de trehaloză, Galactinol și acid L-Malic (19-21 log2FC), în timp ce acidul Oleic, acidul 2-Oxoadipic și acidul Stearic au fost metaboliții cei mai regulați în sus în S2 (15-18 log2FC). În comparația S2 vs. S3, dihidroxiacetona, sulfatul de indoxil și n-Acetilactozamina au fost metaboliți cu cea mai mare creștere în S2 (17-19, 5 log2fc) și trehaloză, Galactinol și acid L-Malic (19-21 log2FC) în S3. În S3 vs. Comparația S4, acidul L-Malic, DL-arginina și acidul Oleic au fost reglate în sus în S3 (16-19 log2fc) și 1,7-Dimetilxantină, D-manoză și n-Acetilactozamină (15-17 log2FC) în S4. În comparația S4 vs. S5, Mononucleotida Flavinei, trehaloza și acidul Izoferulic au fost reglate în sus în metaboliții S4 (18-20 log2FC), în timp ce PG(16:0/18:1(9Z)), D-prolină și DL-arginină au fost reglate în sus (toți metaboliții 16-18 log2FC).
analiza comparativă interspecifică a DEGs în diferite stadii de dezvoltare
Total DEGs în timpul dezvoltării fructelor
comparație interspecifică în etape pereche (LR1 vs.LB1, LR2 vs. LB2 etc.) arată că 928 grade au fost împărțite de toate cele cinci perechi (Fig. 8A). Cel mai mare număr de DEGs a fost identificat în etapa 3 (3989), iar cel mai mic în etapa 4 (2825) (Fig. 8B); întrucât cel mai mare număr de grade unice pentru o pereche au fost observate în etapele 3 (574), 1 și 5 (ambele 554), iar cel mai mic în etapa 4 (126) (Fig. 8A). Numărul de grade reglementate în sus și în jos a fost relativ similar în fiecare dintre comparațiile etapelor pereche; de exemplu, în etapa 5, 1668 grade au fost reglate în sus și 1670 grade au fost reglate în jos în LR în comparație cu LB (Fig. 8B). Cu toate acestea, în celelalte patru etape, numărul de gene reglate în sus a fost ușor (93 până la 189 grade) mai mare.

genele exprimate diferențiat (DEGs) între fructele de L. barbarum (LB) și L. ruthenicum (LR). (A) comparații detaliate în etape (1 până la 5) (LB vs.LR). (B) Numărul de grade superioare (roșii) și inferioare (verzi) în LR în comparație cu LB în cinci etape de dezvoltare studiate.
Transcriptome-pathways
analiza hărții termice a DEGs indică faptul că fructele a două specii prezintă profiluri de expresie genetică foarte diferite în toate etapele de dezvoltare, dar replicile biologice au prezentat profiluri foarte similare, indicând o cantitate limitată de variabilitate individuală în fiecare etapă de dezvoltare (figuri suplimentare: Fig. S9). Analiza comparativă a îmbogățirii căii KEGG arată că doar unele căi au fost în mod constant foarte îmbogățite (În ceea ce privește reglarea genelor) în LR în comparație cu LB pe parcursul tuturor celor cinci etape de dezvoltare (Fig. 9). În special, transducția semnalului hormonal al plantelor (a 2-a cea mai mare în S1, a 8-a cea mai mare în s2, cea mai mare în S3, S4 și S5) și interacțiunea plantă-agent patogen (cea mai mare în S1, a 3-A cea mai mare în S2, A 2-a cea mai mare în S3, a 4-a cea mai mare în S4 și a 15-a cea mai mare în S5) au fost relativ puternic reglate în toate etapele. Biosinteza fenilpropanoidă (nu în top 15 în S1, cea mai mare în S2, a 3-A cea mai mare în S3, a 7-a cea mai mare în S4, a 11-a cea mai mare în s5), ubiquinonă și alte biosinteze terpenoide-chinone (nu în top 15 în S1, a 6-a cea mai mare în S2, A 7-a cea mai mare în S3, a 2-a cea mai mare în S4, a 6-a cea mai mare în S5) au fost, de asemenea, relativ înalt reglate în toate etapele, cu excepția primei. Calea de biosinteză a flavonoidelor nu a fost foarte îmbogățită în stadiile incipiente (nu în top 15 în S1, Al 14-lea cel mai înalt în S2) și foarte îmbogățită în etapele târzii (al 3-lea până la al 4-lea cel mai înalt în etapele 3-5). metabolismul acidului Linoleic (alfa-)a fost foarte îmbogățit în etapele medii (locul 9-Cel mai înalt în S1, locul 2-cel mai înalt în s2, locul 5 și locul 6-cel mai înalt în S3, locul 8-cel mai înalt în S4, nu în top 15 în S5).

analiza comparativă a îmbogățirii căii metabolice KEGG. Primele 15 căi îmbogățite în L. ruthenicum în comparație cu L. barbarum sunt prezentate în stânga (roșu), iar cele îmbogățite în L. barbarum în comparație cu L. ruthenicum în dreapta (Verde). Etapele de dezvoltare (1-5) sunt indicate în figură. valoarea q este o valoare p ajustată FDR.
printre căile reglate în jos în L. ruthenicum în comparație cu L. barbarum (Fig. 9) s-au observat modificări notabile între stadiile incipiente (1 și 2), când metabolismul cianoaminoacidului și biosinteza carotenoidelor au fost cel mai puternic reglate în jos și etapele târzii (4 și 5), când interacțiunile laț în transportul vezicular, metabolismul nicotinatului și nicotinamidei și metabolismul porfirinei și clorofilei au fost constant relativ puternic reglate.
Transcriptome – Gene individuale
dintre genele cele mai exprimate diferențiat, unele au fost specifice stadiului de dezvoltare (adică reglementate foarte diferențiat doar în stadiile de dezvoltare timpurii sau târzii), dar unele au fost reglementate în mod constant foarte diferențiat în toate cele cinci etape studiate (setul de date suplimentare S4). Mai multe gene legate de imunitate foarte puternic reglate în LR în comparație cu LB în stadiile incipiente de dezvoltare se numără printre exemplele modelului de Expresie specific stadiului de dezvoltare: chitinaza a fost a 2-a cea mai mare deg reglată în sus în S1 (13,43 ori), cea mai mare în s2 (13,89 ori), dar în etapele ulterioare nu a fost un DEG. În mod similar, receptorul EIX 1/2 a fost, de asemenea, foarte reglat în stadii incipiente, cel mai mare în S1 (13,70) și al 2-lea cel mai mare în s2 (10,90), dar, de asemenea, nu a fost identificat ca DEG în etapele ulterioare. Unele gene legate de creștere au prezentat, de asemenea, un model de Expresie similar: fosfoglicerat kinaza (PGK) a fost printre puținele gene cu cea mai mare reglare în primele trei etape (13.14, 12.87 și 12.77), dar, de asemenea, nu a fost identificat ca DEG în etapele ulterioare. Ccr4-nu subunitatea complexă de transcripție 7/8 (CNOT7/8) a prezentat, de asemenea, un model de Expresie foarte similar: foarte reglat în primele trei etape și nu un grad în etapele 4 și 5. Mai multe gene asociate biosintezei flavonoide și fenilpropanoide au prezentat un model de Expresie specific stadiului de dezvoltare inversat, cu o expresie relativ scăzută în stadiile incipiente și foarte ridicată în etapele ulterioare. Exemple sunt: dihidroflavonol bifuncțional 4-reductază / flavanonă 4-reductază (DFR), care a fost ușor reglat în LR în S1 (2.25), nu un grad în S2, foarte reglat în sus în S3 (7,79) și a 3-A cea mai mare genă reglată în sus în S4 (14,25) și S5 (16,03). Un Paralog al acestei gene a prezentat un model aproape identic: ușor reglat în sus în S1 (2,44), nu un DEG în s2, foarte reglat în sus în S3 (7,40), al 6-lea cel mai mare DEG reglat în sus în S4 (13,26) și al 5-lea în s5 (14,59). În mod similar, flavonoidul 3′,5′-hidroxilaza (F3’5′ H) nu a fost un grad în primele două etape, foarte reglat în S3 (6,69), a 5-A cea mai mare genă reglată în S4 (13,42) și a 4-a cea mai mare în s5 (15,05). Flavonoid o-metiltransferaza (OMT) nu a fost un DEG în S1, dar în S2 a prezentat deja un nivel mediu-ridicat de reglare în sus (4,32), de către S3 a fost deja al treilea cel mai mare DEG reglat în sus (13,30) și a fost cea mai mare genă reglată în sus în S4 (18,73) și S5 (18,10). Leucoantocianidin dioxigenaza (LDOX; biosinteza antocianinei) nu a fost un grad în S1 și s2, urmată de o reglare ridicată până la foarte mare în etapele ulterioare (5,63, 9,44, respectiv 11,56). Doi paralogi de chalcon sintază (CHS și CHS2; biosinteza flavonoidelor) nu au fost, de asemenea, foarte reglementați în S1 și S2( CHS2: nu a DEG, CHS: -1.14 în S1, nu un DEG în S2), dar în S3–S5 ambele gene au prezentat o reglare medie-înaltă până la înaltă (CHS2: 5.32, 7.84, 6.00; și CHS: 4.67, 7.01, 6.82; respectiv). Am selectat aceste gene pentru analiza qPCR, iar rezultatele sunt foarte congruente cu datele ARN-seq (rezultate suplimentare; Set de date suplimentare S5). În cele din urmă, citokinin dehidrogenaza, o genă legată de biosinteza zeatinei, a fost, de asemenea, din ce în ce mai reglată în ultimele trei etape (2,6-5,7).
cu toate acestea, unele gene au fost exprimate în mod constant diferențiat în toate cele cinci etape studiate. Exemplele au inclus, de asemenea, unele gene legate de imunitate, cum ar fi două paraloguri de glutation S-transferază, foarte reglate în LR în comparație cu LB în toate etapele: 9,38 și 8,58 (toate valorile prezentate ca modificări de pliere în ordinea respectivă) în S1, 6,30 și 6,34 în S2, A 2-a și a 7-a cea mai mare reglare în S3 (14,08 și 12,70), a 2-a și a 4-a cea mai 14.16) și a 2-a și a 6-a cea mai mare în S5 (16.48 și 14.40). Proteina RPM1 de rezistență la boala plantelor a fost, de asemenea, foarte reglată în toate cele cinci etape (S1 = 13,15; s2 = 12,08; S3 = 13,11, S4 = 12,81; S5 = 13:94). Printre genele exprimate în mod constant diferențiat de-a lungul tuturor etapelor de dezvoltare s-au numărat și unele legate de metabolismul aminoacizilor, dar modelul lor a fost inversat în comparație cu exemplele anterioare: au prezentat o reglare descendentă ridicată în LR comparativ cu LB. Exemple sunt acetil-CoA aciltransferaza 1 (aat1; degradarea valinei, leucinei și izoleucinei), cu un profil temporal de reglare descendentă din ce în ce mai mare, începând de la -7,0 în S1 până la <−de 10 ori în ultimele trei etape. Prolina iminopeptidaza, asociată cu metabolismul argininei și prolinei, a fost puternic reglată în LR în toate etapele: S1 = -9,75, s2 = -10,89 (a 3-A cea mai mare), S3 = -11,05 (a 4-a cea mai mare), S4 = -10,01 și S5 = -11,98 (a 3-a cea mai mare). În cele din urmă, 5-metiltetrahidropteroiltriglutamat–homocisteină metiltransferază (metE) a fost constant extrem de scăzută în LR în toate etapele: 2-cea mai mare în S1 (-11,76), cea mai mare în s2 (-11,74), 3-cea mai mare în S3 (-11,43), cea mai mare în S4 (-12,36) și 2-cea mai mare în s5 (-12,83). Două gene asociate replicării ADN și transcripției au fost, de asemenea, foarte reglate în LR în toate etapele: proteina nucleară care leagă GTP Ran (RAN; -10,0 până la -12,0) și factorul de replicare A1 (RFA1) (-8,0 până la -12,0). Unele gene legate de creștere și stres au fost, de asemenea, puternic reglate în mod constant în LR: ribonucleoproteina nucleară eterogenă A1/A3 (hnRNP; -7 până la -11) și proteina de șoc termic 70 kDa 1/8 (HSPA1_8) S1 = -4,95, s2 = -8,88, S3 = -11,48 (a 2-a cea mai mare), S4 = -9,36, S5 = -12,89 (cea mai mare). În mod intrigant, o genă legată de biosinteza fenilpropanoidă, shikimat hidroxicinnamoiltransferaza (HCT), a fost, de asemenea, constant reglată în LR: S1 = -6,82, s2 = -8,14, S3 = -11,71 (cea mai mare), S4 = -11,00 (a 3-A cea mai mare), S5 = -11,91 (a 4-a cea mai mare). Cu toate acestea, un regulator cheie al biosintezei antocianinei, factorul de transcripție MYB114, a fost foarte reglat în LR în toate cele cinci etape de dezvoltare: 6.11, 4.69, 7.47, 9.05 și 8.95 (S1–S5 respectiv).
căi Metabolomice
am efectuat, de asemenea, o analiză comparativă interspecifică etapizată a căilor metabolice (Fig. 10). În prima etapă de dezvoltare (S1), am identificat 39 de metaboliți reglementați diferențiat. Printre primele 20 de căi cu care au fost asociați acești metaboliți, mai multe dintre ele au fost asociate cu aminoacizi, dar numărul total de metaboliți pe cale a fost relativ mic (1-2), iar valorile P nu au sugerat un nivel ridicat de semnificație (Fig. 10-panou 1). În special metabolismul vitaminei B6 (m) și microbian m în diverse medii, ambele au prezentat valori P relativ ridicate, factor de îmbogățire (EF) de 1,0 și 2 metaboliți identificați. În S2, am identificat 58 de metaboliți reglementați diferențiat, asociați cu doar patru căi: triptofan m, biosinteză fenilpropanoidă (b), B de fenilpropanoizi (acestea sunt două căi diferite în baza de date KEGG) și fenilalanină, tirozină și triptofan b (toate EF = 1,0, 2-3 metaboliți și p < 0,5; Fig. 10-panoul 2). În S3, am identificat 59 de metaboliți reglementați diferențiat, asociați cu 19 căi, majoritatea cu EF 1,0, dar valori P relativ nesemnificative (>0,5; Fig. 10-panoul 3). Căile cu un număr relativ mare de metaboliți (n = 5) au fost: digestia și absorbția proteinelor, B de metaboliți secundari din plante, B de antibiotice și b de aminoacizi. În S4, am identificat 58 de metaboliți reglementați diferențiat, asociați cu un număr mare de căi, mai ales cu EF 1,0, și valori de semnificație relativ ridicate (mai ales P > 0,5; Fig. 10-panou 4). Căile cu un număr relativ mare de metaboliți (n-3) au fost: fenilpropanoidul b, fenilalanina, tirozina și triptofanul b, glucozinatul b, B de alcaloizi derivați din calea shikimatului și 2-oxocarboxilicul m. în fructul copt (S5), am identificat 39 de metaboliți reglementați diferențiat, asociați cu un număr mare de căi, dar mai ales cu valori P scăzute și doar 1 metabolit pe cale (Fig. 10-panoul 5). Căile de atac cu mai mult de 1 metabolit au fost: digestia și absorbția proteinelor, fenilpropanoidul b, absorbția mineralelor, carbonul central m în cancer, B al metaboliților secundari, B al fenilpropanoidelor și Arnt aminoacy b. analiza componentelor principale (PCA) a tuturor datelor (2 specii 5 etape 5 x 5 replicate biologice) a evidențiat o similitudine ridicată între replicatele biologice (clustering) și a coroborat variabilitatea notabilă între diferitele etape de maturare a fructelor pentru ambele specii (Fig. 10-panoul 6).

analiza comparativă a îmbogățirii căii metabolice KEGG. Primele 15 căi îmbogățite în L. ruthenicum în comparație cu L. barbarum sunt prezentate în stânga (roșu), iar cele îmbogățite în L. barbarum în comparație cu L. ruthenicum în dreapta (Verde). Etapele de dezvoltare (1-5) sunt indicate în figură. valoarea q este o valoare p ajustată FDR.
metabolom-metaboliți individuali
lista (setul suplimentar de date S6) a metaboliților cu cea mai mare diferențiere reglementată între cele două specii a prezentat unele variații între cele cinci etape de dezvoltare a fructelor. În mod intrigant, fructoza 1-fosfat a fost metabolitul cel mai bine reglat în LR, comparativ cu LB, în toate cele cinci etape: modificarea log2 Fold = 6,3, 7,6, 7,7, 8,1 și 6,5 (respectiv etapele 1 până la 5). În ceea ce privește metaboliții puternic reglați în LB, au existat mai multe variații între etape: în S1, diferențele au fost destul de mici, 9-Decenol fiind metabolitul cel mai bine reglat (modificare de log2 ori = 2,7; comparativ cu LR). Analizele S2 și S3 au produs rezultate foarte congruente, fenolul (3,7 și, respectiv, 3,2) fiind metabolitul cel mai puternic reglat în sus. În S4, sulfatul de indoxil (4.7) a fost principalul metabolit reglat în sus în LB. S-a observat o schimbare metabolică la fructele coapte (S5), unde lista metaboliților reglați în LB a fost completată de stearoilcarnitină (7.1), acid Metoxiacetic (5.3), s-metil-5′ – tioadenozină (4.7), lisinopril (4.7), adenozină 3′, 5′-fosfat ciclic (cAMP) (4.7) etc. Alți metaboliți cu reglaj ridicat în LR (cu excepția fructozei 1-fosfat) au fost naringina (6,2), lauroil-CoA (4,8), L-Fneilalanina (4,6) etc.