Figura 4: structura ochiului.Această imagine este preluată din Chirstie (ref. 4) și este restricționat în funcție de sursa dată (adică nu este opera proprie a autorilor).
structura ochiului: zona din interiorul ochiului (vezi Figura 4) care detectează lumina și culoarea se numește retină. Cele două tipuri de celule de detectare prezente, tije și conuri, procesează informațiile care vin prin lentilă și o trimit pe nervul optic către creier. Celulele de tijă (decare există în jur de 100 de milioane) detectează gradul de ușurință care intră în ochi și sensibilitatea lor depinde de cantitatea de rodopsină prezentă, care este ea însăși generată în interiorul celulelor. Cu toate acestea, rodopsina este distrusă dealbire la expunerea la lumină și, prin urmare, celulele tijei funcționează numai în lumină scăzută asla iluminare ridicată nivelul redus al acestui pigment fotosensibil duce la o sensibilitate foarte scăzută. Celulele conice (dintre care există aproximativ 3 milioane) sunt, de asemenea, sensibile la nivelurile de lumină, dar își păstrează funcția până la o iluminare ridicată prin utilizarea pigmentului iodopsină.Detectarea culorii este o funcție a celor trei tipuri de celule conice prezente în retină: între ele acoperă spectrul vizibil. Acest lucru se datorează faptului că fiecaretip este sensibil la o gamă diferită de lungimi de undă, cu maxime corespunzătoare roșu (lung), verde (mediu) sau albastru (scurt).
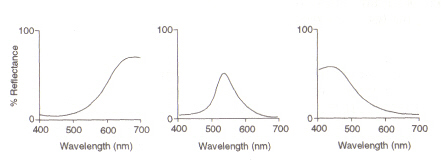
Figura 5: maximele celulelor conice roșii, verzi și albastre (din stânga). Această imagine este preluată din Chirstie (ref. 4) și este restricționat dreptul de autor conform sursei date (adică nu este opera proprie a autorilor).
mecanismul rodopsinei în interiorul celulelor Rodice: proteina rodopsină conține complexul de bază protonat retinal-Schiff, care se află în mod natural în buzunarul inter-membrană format din cei șapte receptori a-helicalici trans-membranari. Există multe discuri plate de rodopsină în segmentul exterior al celulei arod care, la detectarea luminii, suferă o schimbare foto-izomerică de la rodopsină (11-cis) la retină trans-trans. După cascada de fotoizomerizare care are loc prin intermediul a 5 intermediari cu viață scurtă (organigrama 1), trans retina se difuzează și este transformată înapoi în 11-cisretinal înainte de reintrarea în ciclu. Acest proces are loc prin reducerea toall-trans retinol urmată de oxidare/izomerizare în întuneric. Fotoexcitedrodopsina (a 4-a din cei 5 intermediari) declanșează un proces enzimatic în cascadă care are ca rezultat hidroliza GMP. Aceasta, la rândul său, închide canalele specifice cationilor în membrana celulară a tijei, care sunt deschise în mod natural la afluxul de Na+în întuneric și, datorită efectului hiperpolarizării, corpul sinatic intern trimite un semnal nervos către alți neuroni din retină. În cele din urmă, scăderea nivelului de calciu indusă de lumină ajută la recuperarea neuronilor excitați la o stare pasivă, „întunecată”și ciclul începe din nou la detectarea luminii. Fotoreceptorii celulelor conice sunt, de asemenea, șapte a-helicalreceptori cu 11-CIS-retinal ca cromofor. Intervalul de detecție variază de la verde la roșu, deoarece cele trei reziduuri nepolare care conțin hidroxil nearretinal sunt înlocuite secvențial cu cele polare.

stânga-Figura 6: poziția retinei 11-cis în cadrul rodopsinei. Această imagine este preluată din Stryer (ref. 6) și este restricționat dreptul de autor conform sursei date (adică nu este opera proprie a autorilor).
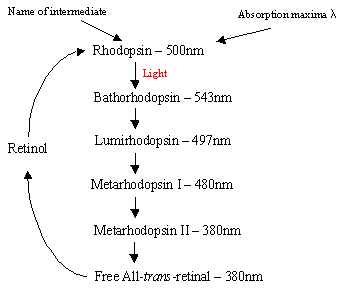
Organigrama Dreapta1: Cei 5 intermediari ai ciclului rodopsinei.
de unde provine rodopsina?
oamenii nu pot producehodopsina, în schimb folosesc și sursa externă, B-carotenul, care se găsește în alimente pentru a-l sintetiza:
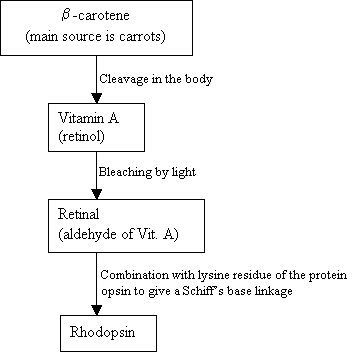 |
Diagrama 2: sinteza rodopsinei imagine nouă =Figura 7: structurile B-carotenului (sus), vitaminei (mijloc) și 11-cis retiniene (jos). |
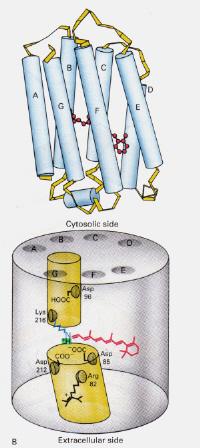
cum diferă mecanismul bacteriorhodopsina Găsită în Halobacterii de cea a rodopsinei găsită încelulele roșii ale ochiului uman?: De data aceasta complexul de bază al retinei protonate-Schiff blochează în mod natural un canal prin membrană altfel format din douăcamere adiacente. Complexul trans protonat donează un proton către Asp-85, care permite apoi ieșirea aceluiași proton în partea extracelulară. Fotoizomerizarea la structura 13-cis permite bazei Schiff să preia un proton din reziduul Asp-96 de pe partea citosolică. După reorientarea formei cis către trans, ciclul de izomerizare și pompare a protonilor continuă.